










My SciELO
 Custom services
Custom servicesServices on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkBiotecnología Aplicada
On-line version ISSN 1027-2852
Biotecnol Apl vol.27 no.3 La Habana July-Sept. 2010
RESEARCH
A pH shift-based procedure to screen fructoligosaccharides fermenting yeast or bacterial Straits
Procedimiento para la pesquisa de levaduras y bacterias fermentadoras de fructoligosacáridos basado en cambios de pH del medio
Enrique R Pérez1, Luis E Trujillo2, Juan G Arrieta2, Heidy Pérez3, María A Brizuela3, Gisselle Trujillo3, Lázaro Hernández2
1 Center for Genetic Engineering and Biotechnology, CIGB Sancti Spíritus, Cuba
2 Center for Genetic Engineering and Biotechnology, CIGB Ave. 31 /158 and 190, Cubanacán, Playa, PO Box 6162, Havana, Cuba
3 Cuban Research Institute for sugarcane derivates, ICIDCA, Havana, Cuba
ABSTRACT
Symbiotic, a combination of short chains-carbohydrates with prebiotic properties e.g. fructooligosaccharides (FOS) and biotherapeutical microorganisms, provides positive health effects and well-being in humans and animals. However; the lack of accurate screening methods hampers the possibility to get new fructo-oligosaccharides (FOS)-fermenting yeast or bacterial strains to design potential symbiotic combinations useful for health and food industries. A valuable screening procedure to visually evaluate pure cultures of bacterial and yeast strains able to ferment FOS in liquid or solid rich media supplemented with a pH indicator is described. Using this procedure, 15 FOS-consuming strains isolated from different sources were successfully assayed using this method. This screening procedure is a useful tool in rapid large-scale detection of potential FOS fermenting-strains.
Keywords: Bromothymol blue, fructoligosaccharides, fructoligosaccharides fermenting microorganisms, prebiotics, probiotics, screening, symbiotic
RESUMEN
Los simbióticos, combinación de carbohidratos de cadenas cortas con propiedades prebióticas e.g. fructooligosacáridos (FOS) con microorganismos bioterapéuticos, proporcionan efectos positivos a la salud de humanos y animales. Sin embargo, la carencia de métodos de pesquisa obstaculiza la detección de nuevas cepas de levadura y bacterias capaces de fermentar FOS para el diseño de nuevos simbióticos con aplicación en la salud animal y humana así como en la industria alimenticia. En este trabajo se describe un valioso procedimiento de pesquisa de cultivos puros de cepas de levaduras y bacterias capaces de fermentar FOS utilizando medios ricos sólidos o líquidos suplementados con un indicador de pH. Utilizando el método propuesto se evaluó satisfactoriamente la capacidad de fermentar FOS de 15 cepas aisladas de diferentes fuentes. Este procedimiento de pesquisa resulta una importante herramienta en la detección rápida de cepas con capacidad potencial para fermentar FOS.
Palabras clave: Bromotimol azul, fructoligosacáridos, microorganismos fermentadores de fructoligosacáridos, prebióticos, prebióticos, pesquisa, simbiótico
INTRODUCTION
Symbiotic provides positive desirable clinical effects in humans and animals (1-3). For this reason the selection of bacterial or yeast strains able to efficiently ferment short chains- carbohydrates with prebiotic properties e.g. fructooligosaccharides (FOS), is a subject of permanent interest for health and food industries.
However; the lack of simple and cost-effective direct screening methods hampers the rapid identification of such important microorganisms. Main targets for FOS consume in the colon are Bifidobacteria and Lactobacilli (1). These intestinal bacteria have shown several positive effects upon human or animals wellbeing (4, 5). Also some yeast strains used for medical therapy consume prebiotics (6).
According to that explained above, if certain carbohydrates, such as fructoligosaccharides, are fermented by only specific beneficial microbial strains, then a liquid or solid growth medium containing these “prebiotic” substrates as unique carbon sources Could efficiently and directly select for those useful FOSfermenters microbial strains.
However, two main drawbacks make it difficult to establish an accurate screening procedure based on FOS consumption. Firstly, FOS commercial preparations in general, may have different polymerization degrees and/or contain contaminants glucose, fructose, sucrose, or other fermentable sugars being rather difficult to establish if microbial growth in FOS-containing medium is really due to FOS metabolism.
This fact makes necessary the use of additional time-consuming confirmatory analytical techniques such as thin layer chromatography (TLC) or High Pressure Liquid Chromatography (HPLC). Secondly, the selection of FOS-fermenter strains by using continuous culture in rich media such as De Man, Rogosa and Sharpe (MRS) (7) to evaluate FOS consumption by bacteria or yeast undoubtedly could mask the final results. To overcome this problem, instead of a rich media like MRS or any similar, the use of minimal medium might be the best choice to evaluate FOS fermentation, but another problem arises.
Contrary to yeasts, Lactobacillus and Bifidobacterium strains for example, need elevated nutritional requirements in the media, and their growth in minimal medium is almost null.
Following this line we developed a new feasible and simple pH shift-based procedure for rapid screening of pure cultures of FOS-fermenting yeast and bacterial strains growing in rich media. As the principle, FOS fermentation by bacteria and yeast provokes acidification of the culture medium (pH below 6) due to production of organic acids. By contrast, microbes unable to metabolize FOS, for example enteric bacteria like Escherichia coli or Salmonella sp., use the nitrogen-containing components in yeast extract or peptone causing alkalinization (pH above 7) due to ammonia release.
The slightly pH switches from acidic to basic conditions can be detected by the addition of the innocuous pH indicator Bromthymol blue (transition interval pH 6.0-7.6; yellow-blue, Sigma Co., USA).
At this stage, a simple visual inspection is enough to detect color change in liquid or solid media allowing a rapid and accurate screening and detection for those microorganisms able to metabolize FOS.
The screening of several potential FOS-consuming strains can be carried out in covered multiwell plastic dishes with 12 or 24 wells (Linbro, Flow Laboratories Inc. USA or Costar, Corning Inc. USA) containing low quantities of the adequate growth medium.
MATERIAL AND METHODS
Microorganisms
Bifidobacterium bifidum 15696, Bifidobacterium dentium 27678 were obtained from the American Type Culture Collection (ATCC) and Escherichia coli TOP10F, Pichia pastoris GS115, Pichia pastoris X33 were purchased from Invitrogen SA.
Other bacterial and yeast strains such as: Saccharomyces boulardii L/25/4/96, Saccharomyces cerevisiae L/25-7-82, Saccharomyces cerevisiae L/25-7-76, Kluyveromyces fragilis L/12-8-1 Kluyveromyces fragilis L/12-8-6, Lactobacillus acidophilus B/103-5, Lactobacillus rhamnosus B/103-1-5, Lactobacillus reuteri B/108-1, Lactobacillus fermentum B/103-11-3, Lactobacillus casei B/103-11-6, Lactobacillus paracasei B/103-11-7, Lactobacillus bulgaricus B/103- 12-6, Streptococcus thermophilus B/103-12-7, used in this research were isolated from different sources and belongs to the microorganism collection of the Cuban Research Institute for Sugarcane Derivates (ICIDCA) Havana, Cuba.
Culture media
The commonly used minimal Yeast Nitrogen base (YNB) and rich media YP and LB (Luria bertani) for yeast and bacterial grow respectively, were prepared according to the Pichia expression vectors for constitutive expression and purification of recombinant proteins Catalog nos. V200-20 and V205-20, Invitrogen SA (USA). Minimal M9 media was purchased from Sigma Co., (USA) catalog number: M6030 and prepared according to the manufactures instructions. Solid media was prepared similar that liquid media but including 20 g of agar per liter. For the pH shift assays using any of the solid or liquid media mentioned above, 0.0025% of Bromothymol blue (transition interval pH 6.0-7.6: yellow-blue, Sigma Co., USA Catalog number: 34656), filter-sterilized solutions of glucose, fructose and sucrose (BDH) added at 2% final concentration or FOS solution obtained in this work 2 and 3% final concentration were incorporated to the adequate culture media according to the different experiments requirements. Final pH in the different media was adjusted to 6.5.
Microbial growth under anaerobic conditions
COY Chambers (COY Laboratory Products Inc.) were used to create and maintain anaerobic conditions as needed during growth of some of the microorganisms mentioned above in the different experiments.
Sugars separation by High Performance Liquid Chromatogrphy (HPLC)
Sugar composition in the enriched FOS solution used in this research were separated in an Aminex HPX- 42C column (0.78 by 30 cm; Bio-Rad Laboratories, Hercules, Calif.) and detected with a RI-410 (Waters) detector. The column temperature during the analysis was kept constant at 85 ºC, and water was used as the mobile phase at a flow rate of 0.6 mL/min.
RESULTS AND DISCUSSION
As the first step of this research we found that unlike yeasts, some lactobacilli and bifidobacteria strains were not able to grow in minimal media supplemented with FOS.
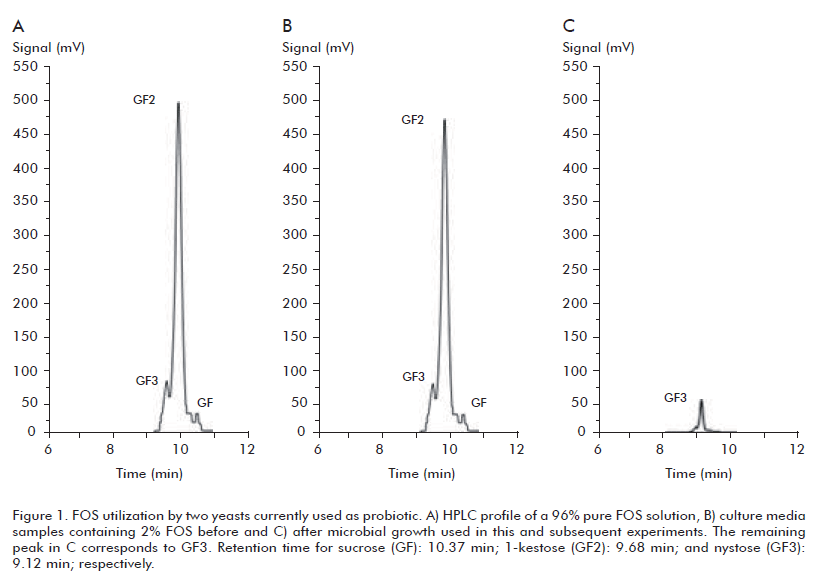
Previously, syrup composed of 4.6% nystose (GF3), 56% 1-kestose (GF2), 21% sucrose (GF), 17% glucose (G), and 1.4% fructose (F) was produced during sucrose transformation by recombinant P. pastoris cells entrapped in Ca-alginate beads. After sugars separation, an enriched FOS solution containing 1-kestose (96%), nystose (3%) and sucrose (1%) was obtained, as judged by HPLC analysis (Figure 1A).
To evaluate microbial FOS fermentation, the use of minimal media appears to be ideal because of the lack of carbohydrates or proteins as alternative energy sources for cell growth. On this basis, the enriched filter-sterilized FOS solution was incorporated to a final concentration of 2 and 3% into solid or liquid minimal Yeast Nitrogen Base (YNB, Invitrogen Co., USA) or minimal M9 (Sigma Co., USA) medium supplemented with 0.05% L-cysteine. Two yeasts (S. boulardii and K. fragilis) and 2 bacterial strains (B. bifidum and B. dentium) were streaked on YNBFOS or M9-FOS plates and incubated 48 h at 30 ºC or 37 ºC respectively, under anaerobic conditions. The assayed yeasts were able to grow in solid or liquid YNB-FOS demonstrating that FOS was equally as good substrate as glucose (BDH), fructose (BDH) and sucrose (BDH) in supporting growth (Figure 1B). Additionally, HPLC analysis confirmed the complete depletion of the FOS fraction after microbial growth (Figure 1C). Unlike yeasts, the two Bifidobacteria and two additional FOS-consumers Lactobacillus strains used as controls of this experiment, failed to grow in solid or liquid M9-FOS (results not shown). Likely their limited growth on minimal media may not guarantee the initial production of enzymes for FOS hydrolysis and transport inside the cell. The fosABCDXE genes encoding for a repressor, a transporter system and a -fructosidases are organized as an operon co-transcribed as a single mRNA in several Lactobacillus and Streptoccocus strains for FOS utilization (8, 9).
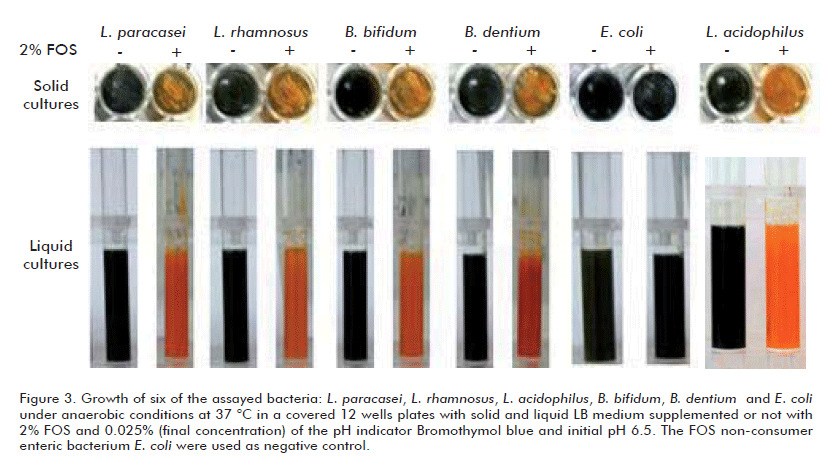
Lactobacilli and Bifidobacteria, organisms that are generally considered to be desirable members of the colonic microbiota, release mainly lactic and acetic acids during FOS catabolism under anaerobic conditions (1) so, provoking acidification of the culture medium reaching pH values below 5. As shown above, these bacteria are unable to grow in minimal media due to their high nutritional requirements. These characteristics prompted us to test whether the addition of an adequate pH indicator as Bromothymol blue (transition interval pH 6.0-7.6: yellow-blue) to the LB media supplemented with 2% FOS could produce visible changes in the medium depending on the bacterium ability to consume FOS.
As shown in table 1 or figures 2A and B, the growth of 15 of the screened bacterial strains, in solid and liquid LB media turned the medium color from initial green (pH 6.5) to yellow indicating acidifiation, pH below 6, due to FOS consumption (Table 1). On the other hand, when FOS was not added, the medium color turned to blue revealing alkalinization due to ammonia release from utilization of the nitrogen-containing carbon sources in yeast extract and peptone so, pH values raised up from initial 6.5 to 8 (Table 1). The same results were obtained when five yeast strains were assayed (Figures 3A and B). Additionally, the use of YNB-FOS minimal medium supplemented with Bromothimol blue revealed also, by simple visual inspection, the capacity to consume these short chain carbohydrates by these yeasts (results not shown).
As expected, the enteric bacterium E. coli and the methylotrophic yeast P. pastoris, which are unable to use FOS, turned the medium color to blue raising the pH values about 8 (Table 1) due to also utilization of the nitrogen-containing carbon sources in yeast extract and peptone.
FOS depletion was further verified by HPLC analysis demonstrating that the assayed bacteria and yeasts consumed totally the GF2 (1-kestose) and GF3 (nystose) fractions (results not shown). In addition, no accumulation of other monosaccharide was seen which suggest that they were also degraded by the microorganisms. These results agree with previous reports dealing with efficient consume of GF2, GF3 together with mono and disaccharide fractions (10).
It has been established that the diet, host phylogeny and gut morphology influence the microbial ecology of the gastrointestinal tract (11,12) The beneficial potential of lactic acid bacteria and yeast has been shown to be enhanced by the introduction of prebiotic carbohydrates mainly of fructo- and galactooligosaccharidic nature (6). In recent studies other authors concluded that prebiotics selectively enhance lactobacilli and bifidobacteria populations and reduce colonization by pathogenic bacteria (13-15).
However, there are not reports regarding to rapid and accurate qualitative detection methods to get new potential fructoligosaccharides-fermenting yeast or bacterial strains based on consume of these short chain carbohydrates widely used as prebiotics. Previous studies have been aimed to apply in vitro methods for selection of suitable bacterial and yeast strains for human or animal health improvement (16, 17). In this particular report the addition of an innocuous pH indicator like Bromothymol blue (transition interval pH 6.0-7.6: yellow-blue) to any of the used media supplemented with 2% FOS produced rapid and visible changes in the medium color depending on the bacterium or yeast ability to consume or not FOS as unique carbon source. This pH indicator has not previously been used for this purpose. On the other hand the enriched 96% FOS preparation used in this work composed mainly by GF2 (1-kestose) and GF3 (nystose) is useful as a carbon source since guarantee that microbial growth is mainly due to FOS consume as unique carbon source.
CONCLUSION
According to the results described above, the proposed screening method is a valuable tool in large-scale detection of pure cultures of potential FOS fermenters- yeast or bacterial strains for further symbiotic designs.
ACKNOWLEDGMENTS
This research was partially funding by the research project 3031254 of the Center for Genetic Engineering and Biotechnology, Havana, Cuba.
REFERENCES
1. Collins MD, Gibson GR. Probiotics, prebiotics, and symbiotic: approaches for modulating the microbial ecology of the gut. Am J Clin Nutr 1999;69:1052S-7S.
2. Kohler H, McCormick BA, Walker WA. Bacterial-enterocyte crosstalk: cellular mechanisms in health and disease. J Pediatr Gastr Nutr 2003;36:175-85.
3. Patterson JA. Prebiotic Feed Additives: Rationale and Use in Pigs. Adv Pork Prod 2005;16:149-58.
4. Kaplan H, Hutkins RW. Metabolism of fructooligosaccharides by Lactobacillus paracasei 1195. Appl Environ Microb 2003;69:2217-22.
5. Gaggìa F, Mattarelli P, Biavati B. Probiotics and prebiotics in animal feeding for safe food production. Int J Food Microbiol 2010; 141 Suppl 1: s15-s28.
6. Mitterdorfer G, Kneifel W, Viernstein H. Utilization of prebiotic carbohydrates by yeasts of therapeutic relevance. Lett Appl Microbiol. 2001;33:251-55.
7. Sghir A, Chow JM, Mackie RI. Continuous culture selection of Bifidobacteria and lactobacilli from human faecal samples using fructooligosaccharide as selective substrate. J Appl Microbiol 1998; 85:769-77.
8. Goh YJ, Zhang C, Benson AK, Schlegel V, Lee JH, Hutkins RW. Identification of a putative operon involved in fructooligosaccharide utilization by Lactobacillus paracasei. Appl Environ Microbiol 2006;72 (12):7518-30.
9. Goh YJ, Lee JH, Hutkins RW. Functional analysis of the fructooligosaccharide utilization operon in Lactobacillus paracasei 1195. Appl Environ Microbiol 2007; 73(18):5716-24.
10. Saulnier DM, Molenaar D, de Vos WM, Gibson GR, Kolida S. Identification of prebiotic fructooligosaccharide metabolism in Lactobacillus plantarum. Appl Environ Microbiol 2007;73:1753-65.
11. Ley RE, Hamady M, Lozupone C, Turnbaugh PJ, Ramey RR, Bircher JS, et al. Evolution of mammals and their gut microbes. Science 2008;320:1647-51.
12. Ley RE, Lozupone CA, Hamady M, Knight R, Gordon JI. Worlds within worlds: evolution of the vertebrate gut microbiota. Nat Rev Microbial 2009;6:776-88.
13. Biggs P, Parsons CM. The effects of grobiotic- P on growth performance, nutrient digestibilities, and cecal microbial populations in young chicks. Poult Sci 2008; 87:1796-803.
14. Baurhoo B, Letellier A, Zhao X, Ruiz- Feria CA. Cecal populations of Lactobacilli and Bifidobacteria and Escherichia coli after in vivo Escherichia coli challenge in birds fed diets with purified lignin or mannanoligo-saccharides. Int J Poult Scie 2007;86:2509-16.
15. Baurhoo B, Goldflus F, Zhao X. Purified cell wall of Saccharomyces cerevisiae increases protection against intestinal pathogens in broiler chickens. Poult Sci 2009;8:133-7.
16. Ten LN, Im WT, Aslam Z, Larina L, Lee ST. Novel insoluble dye-labeled substrates for screening inulin-degrading microorganisms. J Microbiol Methods 2007; 69(2):353-7.
17. Modesto M, D'Aimmo MR, Stefanini I, Trevisi P, De Filippi S, Casini L , et al. A novel strategy to select Bifidobacterium strains and prebiotics as naturalgrowth promoters in newly weaned pigs. Livestock Sci 2009;122:248-58.
Received in March, 2010.
Accepted for publication in June, 2010.
Luis E Trujillo, Center for Genetic Engineering and Biotechnology, CIGB Ave. 31 /158 and 190, Cubanacán, Playa, PO Box 6162, Havana, Cuba. E-mail: luis.trujillo@cigb.edu.cu

{kind=link}
{kind=link}
{kind=link}
{kind=link}