










My SciELO
 Custom services
Custom servicesServices on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkBiotecnología Aplicada
On-line version ISSN 1027-2852
Biotecnol Apl vol.30 no.3 La Habana July-Sept. 2013
REPORT
Molecular mechanisms involved in the inhibition of tumor cells proliferation exposed to elevated concentrations of the epidermal growth factor
Mecanismos moleculares que participan en la inhibición de la proliferación de células tumorales expuestas a concentraciones elevadas del Factor de Crecimiento Epidérmico
Isabel A Guillén, Jorge Berlanga, Hanlet Camacho, María E Fernández-de-Cossio, Lincidio Pérez, Lidia I Novoa, Daniel O Palenzuela, Tamara Díaz, Gerardo E Guillén, Luis Herrera, Karelia Cosme, Laritza Gorovaya, Osmani Mendoza, Julio R Fernández, Julio A Ancizar, Jose Suarez, Ángela D Tuero, María E Ochagavía, Juan Roca, Jorge Gavilondo, Diana García del Barco, Jorge Martín
Centro de Ingeniería Genética y Biotecnología, CIGB. Ave. 31 e/ 158 y 190, Cubanacán, Playa, CP 11600, La Habana, Cuba.
ABSTRACT
The EGF promotes inhibition of cell proliferation in vitro and in vivo models depending on its concentration, application schema and the type of tumor cells on which it acts. Our research hypothesis was based on the fact that the EGF varies the expression of genes involved in a negative regulation of tumor cell lines proliferation carrying high levels of its receptor (EGFR). Our objectives were, to obtain information about the effect of EGF on tumor cell proliferation in vitro and in vivo models and, know the gene expression patterns of a group of genes involved in cancer signaling pathways and EGFR. The results showed that EGF at nanomolar concentrations inhibits the tumor cells proliferation bearing high levels of EGFR and, promotes the survival of treated animals, establishing a direct relationship between the inhibition of cell proliferation, high concentrations of EGF and, high amount of EGFR in the cells. The differential gene expression profile showed a variation in a group of genes which exert a powerful control over the cell cycle progression, gene transcription and apoptosis. It was concluded that the inhibition of tumor cell proliferation by the action of EGF is due to activation of molecular mechanisms controlling cell cycle progression. This work won the Annual Award of the Cuban Academy of Sciences in 2012.
Keywords: epidermal growth factor, cancer, gene expression, animal model.
RESUMEN
El factor de crecimiento epidérmico (FCE) promueve la inhibición de la proliferación celular en modelos in vitro e in vivo en dependencia de su concentración, el esquema de aplicación y el tipo de células tumorales sobre las que actúa. Nuestra hipótesis de investigación se basó en que el FCE varía la expresión de los genes involucrados en la regulación negativa de la proliferación de líneas celulares de origen tumoral que portan altas concentraciones de su receptor (RFCE). Los objetivos fueron obtener información acerca del efecto del FCE en la proliferación de células tumorales en modelos in vitro e in vivo, y conocer los patrones de expresión génica de un grupo de genes involucrados en las vías de señalización del cáncer y el FCE. Los resultados mostraron que el FCE a concentraciones nanomolares inhibe la proliferación de células de origen tumoral que poseen elevados niveles de RFCE, favorece la sobrevida de los animales tratados, y reduce el índice mitótico de las células del tumor, por lo que se establece una relación entre la inhibición de la proliferación celular, las altas concentraciones de FCE y la elevada cantidad de RFCE en las células. La expresión diferencial de genes reveló la variación en la expresión de genes que ejercen un control potente sobre la progresión del ciclo celular, la transcripción génica y la apoptosis. Se concluyó que la inhibición de la proliferación de las células tumorales por la acción del FCE se debe a la activación de mecanismos moleculares de control de la progresión del ciclo celular. Este trabajo recibió el Premio Anual de la Academia de Ciencias de Cuba en 2012.
Palabras clave: factor de crecimiento epidérmico, cáncer, expresión de genes, modelo animal.
INTRODUCTION
Epidermal growth factor (EGF) is a 6 kDa polypeptide that binds to its receptor (EGFR), a tyrosine kinase that activates transduction signals promoting the transcription of genes involved in growth and proliferation of different cell types. The exogenous administration of supraphysiological concentrations of EGF triggers reparative and cytoprotective effects which lead to cell survival and the repair of internal and external organs [1]. EGF and its receptor have also been implicated in human malignancies because of the interconnection of wound healing and the development of tumors favored by the dynamic interaction of a variety of cell types to promote cell proliferation and survival. Recent evidence from animal models suggests that EGF does not initiate cell transformation, and its role in carcinogenesis is only as a promoting agent, acting as an epigenetic factor when cells are prone to malignancy [2]. It has been observed that depending on EGF concentrations, the application schema and type of cells on which it acts can inhibit or promote cell growth, showing a biphasic action on tumor cell lines of epithelial origin. For example, picomolar concentrations (3-100 pM) of EGF promote tumor cell proliferation, and nanomolar concentrations (0.1-10 nM) cause its inhibition [3-7].
This paradox suggests that the high availability of EGFR in cells and high EGF concentrations produces a negative cooperation between the EGFR dimerized receptors, supporting the formation of a low affinity receptor for the ligand, and activating signaling pathways that have a negative effect on cell proliferation [8].
Taking all these into account, we set out to find genes involved in the negative regulation of the proliferation of tumor cell lines carrying high concentrations of EGFR by exposing them to nanomolar concentrations of the ligand (EGF). For this purpose, we developed the following objectives: to obtain information about the effect of EGF on the proliferation and development of tumor cells in models in vitro and in vivo; and, to know the gene expression patterns of a selected group of genes related to the signaling cancer pathways and EGF.
MATERIALS AND METHODS
We used six cancer cell lines (K562, MCF-7, U1906, LS-174T, H125 and A431) with different concentrations of EGFR. These cell lines were exposed for 72 h to the ligand effect (EGF). An animal model was obtained in nude mice xenografts with the cell line A431 over-expressing the levels of EGFR. The animals were treated subcutaneously in the dorsal region, near to the tumor with 1 µg/kg of EGF. The real time polymerase chain reaction (RT-PCR) technique was used to explore the differential expression of 44 genes involved in EGF signaling pathway and cancer in cells treated with EGF and controls, in in vitro and in vivo models. The quantitative RT-PCR data analysis was performed by RT-Cycler CapitalBio series analysis software, version 2.001 (CapitalBio Co., Ltd., Beijing, China). Relative quantification of gene expression was performed using the software REST 2009 v2.0.13 (Qiagen GmbH, Germany). The LingReg software (version 11.3, 2009, Amsterdam, Netherlands) was used to estimate the efficiency of PCR and, the geNorm software was used for the selection of more stable reference genes [9].
RESULTS AND DISCUSSION
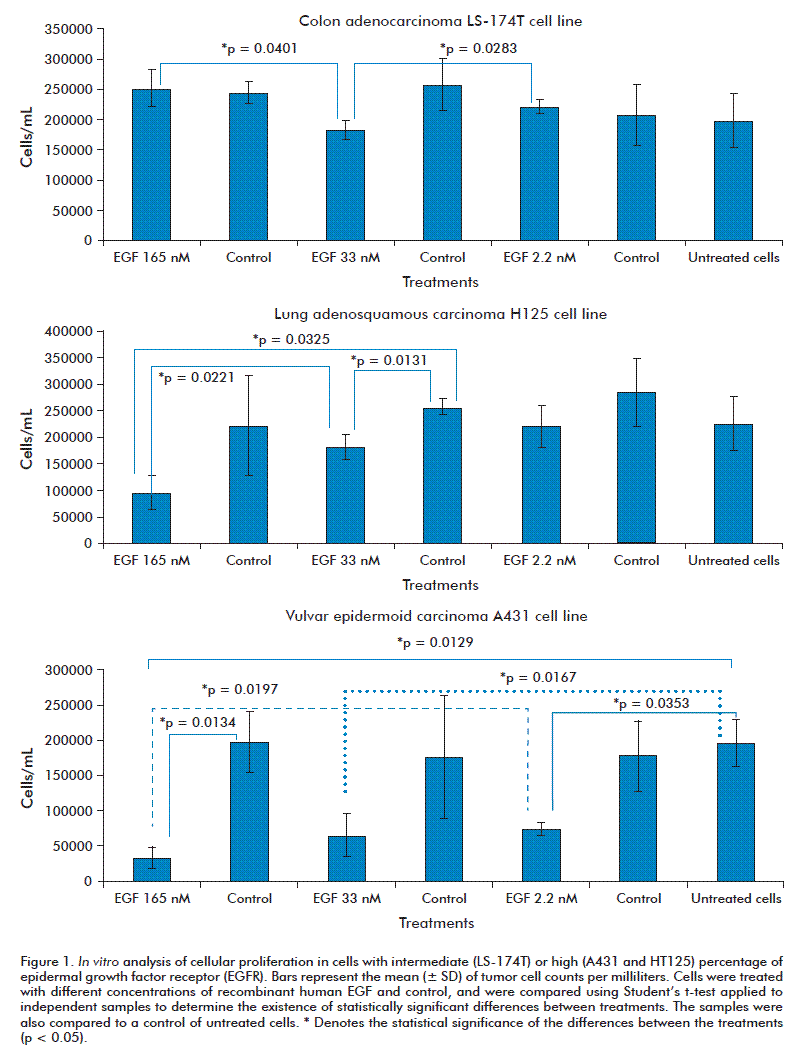
EGF at concentrations of 2.2, 33 and 165 nM does not favor the proliferation of tumor cells. Interestingly was observed that EGF inhibited the proliferation of tumor cells over-expressing the growth factor receptor (EGFR), such as H125 (lung adenosquamous carcinoma cell line) and A431 (epidermoid carcinoma of the vulva cell line). According to these results, there is a direct relationship between the inhibition of cell proliferation, the exposition of cancer cells to high concentrations of exogenous EGF and, the high amount of endogenous EGFR in cells (Figure 1).
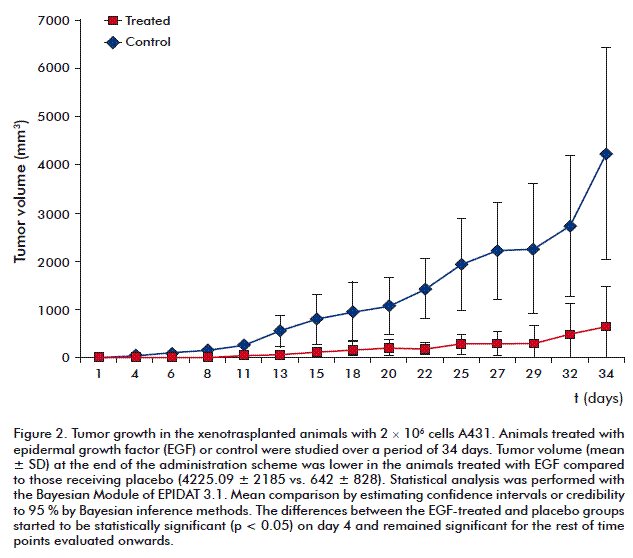
We observed an inhibition of tumor growth in the in vivo model of nude mice xenografted with the A431 cell line and treated with EGF for 5 weeks (Figure 2), there was obtained a decrease in the mitotic index of tumor cells, which is a critical parameter in the prevention and control of cancer [10]. Since EGF may provide protective effects against the aggressiveness and invasiveness of certain types of tumors, it was observed in the in vivo study a significant increase in survival of the animals inoculated with 3 × 106 A431 cells and treated with EGF in comparison with the untreated animals [10]. The EGF favored the survival of animals, perhaps by providing protective properties against tumor aggressiveness and invasiveness. As proposed by Amagase et al. in 1990, EGF plays an important role in preventing metastasis of certain malignancies, thus prolonging the survival time of xenotransplanted athymic animals [11].
The differential gene expression analysis in in vivo and in vitro studies revealed the over-expression of genes that directly and potently regulate cell cycle progression and apoptosis. This was detected by analyzing the differential gene expression profile in the tumors and cell lines.
For the cell line H125, 21 genes varied significantly their expression, with 27 in A431. However, in the U1906 cell line, only 3 genes varied its expression. This shows a correspondence between the magnitude of the gene expression variability and the observed phenotype.
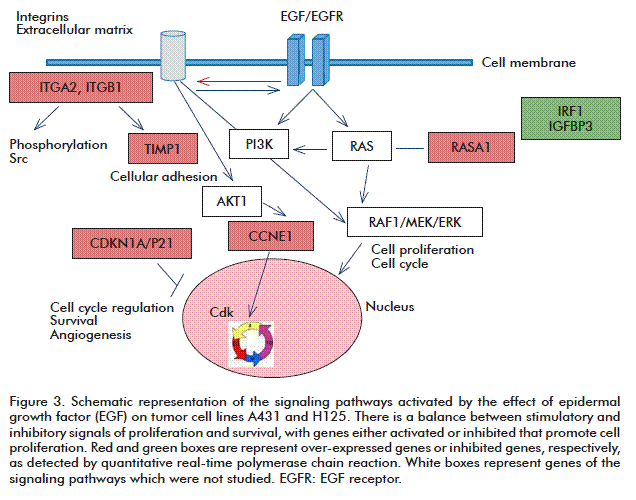
In the A431 and H125 cell lines, similar genes were over-expressed and inhibited; among them having a fold change above 2 (FC ≥ 2): ITGA2, CDKN1A/P21, CCNE1, TIMP1, RASA1, ITGB1, IRF1, and IGFBP3. Most of them are part of the EGF/EGFR signaling cascade, which regulates cell cycle progression and apoptosis.
A scheme of the identified molecular interactions showing the most relevant genes in gene expression analysis is shown in Figure 3. As can be observed there were genes with FC > 1.5, with statistically significant differences between treated and control cells of A431 and H125 tumor cell lines.
Also, a specific group of genes for each cell line were involved in the process of cell proliferation inhibition. In the cell line A431, the activation of the transcription factor STAT1 induces the expression of genes involved in apoptosis and in the inhibition of the TP53 gene, which carries a gain-of-function mutation favoring cell tumorigenicity. Furthermore, in the H125 cell line was observed the over-expression of pro-apoptotic genes, such as caspases 8 and 9.
In general, the molecular mechanisms involved in the inhibition of tumor cell proliferation in both scenarios (in vivo and in vitro), were directed by the variation in the expression of genes that control cell cycle progression, such as the cyclin-dependent kinase inhibitor CDKN1A, and cyclin-dependent kinase 4 (Cdk4). The way in which these genes changed its expression may justify the phenotypic effect mediated by the action of EGF in tumor cells.
As already mentioned, the gene expression for the tumor suppressor gene TP53 varied similarly in the A431 in studies in vivo and in vitro. The TP53 protein controls the cell cycle progression to repair the damage in the genome. In the A431 cell line, TP53 gene has a point mutation p.Arg273His, which is a change at the arginine residue 273 to histidine. This is a gain-of-function type I mutation (type I missense mutation), which affects the binding sites of the p53 protein to the DNA. Therefore, the G1/S checkpoint in the cell cycle does not function properly to repair the genetic damage [12]. This causes a genomic instability and malignant progression of cells. Among the phenotypic effects of the mutated TP53 on the cells are: increased cell proliferation, increased growth density and induction of antiapoptotic activity [13].
The inhibition of the mutated TP53 gene expression in the A431 cell line treated with EGF may be associated to a decrease in tumorigenicity of the mutated protein. A decrease in the levels of this protein may allow the cell to recover somewhat the regulation of cell cycle progression at the G1-S transition, by decreasing Cdk4 levels. We observed a possible link between the effect of EGF and the inhibition of two important cell cycle regulators, the TP53 and Cdk4, and its implications in the inhibition of cell cycle progression from G1 to S phase, thereby, decreasing tumor growth. These results suggest that EGF promotes similar mechanisms in both biological scenarios (in vivo and in vitro) in the A431 cell line, resulting in an inhibition of cell proliferation.
Caspase 9 is another gene in the regulatory cascade of EGF on tumor cells bearing high levels of EGFR. This gene is a member of the aspartic acid protease-cysteine family. The sequential activation of caspases plays a central role in cellular apoptosis, so that the action of EGF on restoring antitumor cell biological pathways may also be taking place through activation of the caspase pathway. The activation of caspases 8 and 9 were also detected in the H125 cell line treated with EGF. This is another regulatory pathway activated in cancer models in vivo and in vitro treated with the EGF. The in vivo scenario promotes transient recovery of the cellular tumor suppressor ability, by reducing gene expression levels of Cdk4 and TP53, and increasing caspase 9 levels. The in vitro models revealed a group of genes that varies in a similar fashion in the A431 and H125 cell lines, which inhibited the EGF-mediated proliferation.
All these genes are involved in the signaling cascade of EGF/EGFR and cancer. It is relevant the activation by EGF of the expression of integrins like ITGA2 and ITGB1 (Figure 3), which in turn activates the signaling cascade through EGFR. These signals may induce the expression of integrins, a family of transmembrane glycoprotein receptors that mediate cell-cell and cell-matrix interactions, playing a fundamental role in cellular events such as proliferation, differentiation and apoptosis. It is also known that the over-expression of the beta1 integrin inhibits cell proliferation through the activation of the cyclin dependent kinase inhibitor p21. Integrin over-expression can reduce cell adhesion and can inactivate the phosphoinositide 3 kinase (PI3K) [14].
The protein encoded by the p21 gene is the cyclin dependent kinase inhibitor 1A (CDKN1A), which regulates the cell cycle S phase progression. The P21 expression is regulated by the tumor suppressor gene TP53. It has also shown to be regulated by other genes such as the integrins, through its effect on the PI3K/AKT1 pathway of the EGFR signaling cascade.
The STAT1 transcription factor is another interesting gene, which, as has been reported by other groups, is activated in the A431 cell line [15]. Its activation is associated with a decrease in cell proliferation and increased apoptosis, and is further required to inhibit the proto-oncogene MYC, which directs cell proliferation through the over-expression of cyclins and gene expression inhibition of p21. In the A431 cell line, the expression of MYC was inhibited, and, therefore, it must favor the cell proliferation inhibition observed in these cells.
On the other hand, the TIMP1 gene is a metalloprotease inhibitor that was over-expressed in A431 and H125 cell lines. This factor forms complexes with matrix metalloproteinases (MMP) and irreversibly inactivates collagenase. MMPs belong to the extracellular matrix (ECM) which activates the integrins. It is known that there must be a balance between the levels of proteases and their inhibitors to form a correct ECM, which is a component of the granulation tissue, a key component of wound healing and tissue regeneration. Moreover, one of the key events in malignant tumor development is the degradation of the ECM, which at the time is essential for angiogenesis and the formation of blood vessels from preexisting blood networks.
Another interesting gene was RASA1, showing increased expression in both, the A431 and H125 cell lines. The RASA1 protein is a negative regulator of the RAS/MAPK signaling pathway, which transmits signals from outside the cell into the nucleus. The signaling pathway through RAS/MAPK directs important functions such as cell growth and division. When RASA1 is mutated, there is a path of deregulation through RAS/MAPK pathway, which promotes uncontrolled proliferation and cancer development.
In this scenario, not everything is an antiproliferative and pro-apoptotic stage, as has been arising from the explanation for the inhibition seen in cell proliferation and tumor growth, promoted by genes such as: TIMP1, p21, Cdk4, ITGA2, ITGB, TP53, MYC, RASA1 and caspases 8 and 9. There was also a group of genes that varied their expression, which participate in cell proliferation promotion. This is the case of the over-expression of the CCNE1 gene, a member of the cyclins family, which positively regulates the G1/S transition.
In a complex scenario such as the EGF/EGFR signaling cascade, it is significant that the cell promotes activation of powerful regulatory mechanisms. Those mechanisms maintain the control over the oncogenic effects that can trigger and support uncontrolled cell proliferation and cancer development.
Recent reports suggest that EGF induces more changes in cell proliferation and death in A431 cancer cells than the anticancer drugs cetuximab or gefitinib [16]. Cell arrest and death events produced by EGF appear to remain for periods longer than those caused by some conventional antitumor drugs. This is important to continue unraveling the complex molecular mechanisms of EGF in cell proliferation inhibition in cancer cells.
CONCLUSIONS
This is the first study in Cuba exploring the mechanism of action of EGF using the RT-PCR technique to quantify the differential gene expression profile of 44 genes involved in cancer pathways and the EGF/EGFR signaling. It is also the first time that the inhibitory effect of EGF on cell proliferation is related to the gene expression levels of the mutant TP53p.Arg273His and the inhibition of cell proliferation in an in vivo and in vitro model using the vulva cancer cell line A431.
The antiproliferative effects of EGF were mediated by changes detected in the expression of genes which regulate cell cycle progression, apoptosis and transcription. They included the effect of the gene dependent kinases inhibitor (CDKN1A) which regulates cell cycle progression, and the caspase 8 and 9, and the transcription factor STAT1 genes.
There is a functional convergence between EGFR expression levels in cells and the signaling through integrins, thus favoring the over-expression of the cell cycle inhibitor gene CDKN1A/P21. It was also observed that EGF at a 2.2 nM concentration significantly promotes survival of treated animals when placed subcutaneously near to the tumor generated in nude mice xenografted with the A431 cell line.
REFERENCES
1. Cross M, Dexter TM. Growth factors in development, transformation, and tumorigenesis. Cell. 1991;64(2):271-80.
2. Berlanga J, Álvarez S, De la Fuente J, López P. Considerations on the transforming potential of the epidermal growth factor. Biotecnol Apl. 1998;15(2):65-9.
3. Barnes DW. Epidermal growth factor inhibits growth of A431 human epidermoid carcinoma in serum-free cell culture. J Cell Biol. 1982;93:1-4.
4. Kawamoto T, Sato JD, Le A, Polikoff J, Sato GH and Mendelsohn J. Growth stimulation of A431 cells by epidermal growth factor: identification of high-affinity receptors for epidermal growth factor by an anti-receptor monoclonal antibody. Proc Natl Acad Sci USA. 1983;80:1337-41.
5. Kawamoto T, Mendelsohn J, Le A, Sato GH, Lazar CS, Gill GN. Relation of epidermal growth factor receptor concentration to growth of human epidermoid carcinoma A431 cells. J Biol Chem. 1984;259:7761-6.
6. Fan Z, Lu Y, Wu X, DeBlasio A, Koff A, Mendelsohn J. Prolonged induction of p21cip1/WAF1/CDK2/PCNA complex by epidermal growth factor receptor activation mediates ligand-induced A431 cell growth inhibition. J Cell Biochem. 1995;131:235-24.
7. Choi J, Moon SY, Hong JP, Song Jy, Oh KT, Lee SW. Epidermal growth factor induces cell death in the absence of overexpressed epidermal growth factor receptor and erbb2 in various human cancer cell lines. Cancer Invest. 2010;28:505-14.
8. Defize LHK, Boonstra J, Meisenhelder J, Kruijer W, Tertoolen LGJ, Tilly BC. Signal transduction by epidermal growth factor occurs through the subclass of high affinity receptors. J Cell Biol. 1989;109:2495-507.
9. Vandesompele J, De Preter K, Pattyn F, Poppe B, Van Roy N, De Paepe A, et al. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002;3(7):RESEARCH0034.
10. Guillén IA, Camacho H, Fernández ME, Palenzuela DO, Pérez L, Ochagavia ME, et al. Effect of human epidermal growth factor on the tumor cell line A431: in vivo analysis of tumor growth inhibition and gene expression. Biotecnol Apl. 2012;29:155-61.
11. Amagase H, Tamura K, Okuhira M, Kakimoto M, Amano H, Hashimoto K, et al. Epidermal growth factor prolongs survival time of tumor-bearing mice. Jpn J Cancer. Res. 1990;81(5):495-500.
12. Williams AC, Miller JC, Collard TJ, Bracey TS, Cosulich S, Paraskeva C. Mutant p53 is not fully dominant over endogenous wild type p53 in a colorectal adenoma cell line as demonstrated by induction of MDM2 protein and retention of a p53 dependent G1 arrest after gamma irradiation. Oncogene. 1995;11(1):141-9.
13. Prasad KA, Church JG. EGF effects on p53 in MDA-468 human breast cancer cells: implications for G1 arrest. Cell Prolif. 1997;30(2):81-94.
14. Banno A, Ginsberg MH. Integrin activation. Biochem Soc Trans. 2008;36(Pt 2):229-34.
15. Gulli LF, Palmer KC, Chen YQ, Reddy KB. Epidermal growth factor-induced apoptosis in A431 cells can be reversed by reducing the tyrosine kinase activity. Cell Growth Differ. 1996;7(2):173-8.
16. Song JY, Lee SW, Hong JP, Chang SE, Choe H, Choi J. Epidermal growth factor competes with EGF receptor inhibitors to induce cell death in EGFR-overexpressing tumor cells. Cancer Lett. 2009;283(2):135-42.
Isabel A Guillén. Centro de Ingeniería Genética y Biotecnología, CIGB. Ave. 31 e/ 158 y 190, Cubanacán, Playa, CP 11600, La Habana, Cuba. E-mail: isabel.guillen@cigb.edu.cu.


{kind=link}
{kind=link}
{kind=link}