










My SciELO
 Custom services
Custom servicesServices on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkBiotecnología Aplicada
On-line version ISSN 1027-2852
Biotecnol Apl vol.31 no.1 La Habana Jan.-Mar. 2014
RESEARCH
Yield improvement of the sea lice MY32/Cr novel antigen production and IgM immune response characterization in Oreochromis niloticus as a model
Rendimiento incrementado del nuevo antígeno MY32/Cr del piojo de mar y caracterización de la respuesta inmune de IgM en Oreochromis niloticus
Liliana Basabe1, Yamila Carpio1, Diamilé Gonzalez2, Antonio Morales1, Mario P Estrada1
1 Departamento de Biotecnología Animal, Dirección de Investigaciones Agropecuarias, Centro de Ingeniería Genética y Biotecnología, CIGB. Ave. 31 e/ 158 y 190, Cubanacán, Playa, CP 11600, La Habana, Cuba.
2 Dirección de Desarrollo Tecnológico, CIGB. La Habana, Cuba.
ABSTRACT
Sea lice cause over 300 million € annual losses in salmon aquaculture-leading countries and most treatments consist of chemicals, with their limitations regarding the generation of parasite resistance, environmental damage and high costs. Researches about fish vaccines against sea lice as a method for controlling parasites have been accomplished; however there is no commercial vaccine available up to date against this pest. It could be due to an insufficient immunological and protective response of the selected antigen or to the lack of appropriate/suitable process for its production with commercial perspectives. Previously, the gene coding for the akirin protein of Caligus rogercresseyi (MY32/Cr) was cloned. It was produced recombinant in Escherichia coli and it showed 57 % of protection in a vaccination-challenge experiment against sea lice infestation in salmon. In the present study, the MY32/Cr yield was improved up to four fold in fermentation process by testing different culture media. It was also studied the effect of this novel antigen, for a potential vaccine, on humoral immune response of tilapia (Oreochromis niloticus). It was observed that the immunization with MY32/Cr elicited a statistically significant IgM antibody response after a double injection of the recombinant protein. The process obtained is a suitable method, an essential tool as a first step to produce this recombinant antigen at commercial scale. The results suggest that the MY32/Cr protein may be used as a target to be tested in salmon, for developing of a vaccine to control sea lice infestations in fish; and that the administration of a single booster dose could be useful to improve antibody response.
Keywords: fish, heterologous expression, IgM, immunization, MY32, sea lice.
RESUMEN
Las pérdidas anuales provocadas por el piojo de mar en los países líderes en la acuicultura salmonera superan los 300 millones de euros. La mayoría de los tratamientos emplean compuestos químicos, con sus limitaciones de generación de resistencia en los parásitos, el potencial daño al medioambiente y sus altos costos. A pesar de los estudios en peces sobre vacunas contra esta plaga como un posible método de control, no existe actualmente ninguna vacuna comercial. Esto pudiera deberse a que los candidatos estudiados no han mostrado un respuesta inmunológica y protectora suficiente, o a la carencia de un proceso apropiado para su producción con perspectivas comerciales. Previamente, se clonó el gen que codifica para la proteína akirin de Caligus rogercresseyi (MY32/Cr). Esta se produjo de forma recombinante en Escherichia coli, y mostró un 57 % de protección contra la infestación por piojo de mar en un estudio de vacunación-reto en salmón. En el presente trabajo se mejoró la productividad de MY32/Cr hasta cuatro veces mediante procesos de fermentación empleando diferentes medios de cultivo. También se estudió el efecto de este nuevo antígeno como preparado vacunal sobre la respuesta inmune humoral en tilapia (Oreochromis niloticus). La inmunización provocó una respuesta de anticuerpos IgM estadísticamente significativa después de la reinmunización con MY32/Cr. El proceso obtenido constituye un método eficiente como premisa esencial para la producción de este antígeno recombinante a escala comercial. Los resultados sugieren que la proteína MY32/Cr podría ser empleada como antígeno blanco para evaluaciones en salmón, con vistas a desarrollar una vacuna para el control de las infestaciones por piojo de mar en peces. La administración de una dosis de reinmunización resultaría útil para mejorar la respuesta inmune mediada por anticuerpos.
Palabras clave: peces, expresión heteróloga, IgM, inmunización, MY32, piojo de mar.
INTRODUCTION
Sea lice are the major pest in the salmon industry causing annual losses over 300 million € [1]. However there is no commercial vaccine available up to date against sea lice. Akirins are among the potential targets for vaccines candidates’ development against this pest.
Akirins affect the expression of signal transduction and innate immune response genes in vertebrates and invertebrates [2]. Recently, the gene encoding the akirin from Caligus rogercresseyi (my32) was cloned to produce the recombinant protein (MY32/Cr) in Escherichia coli [3]. Experiments with recombinant akirins have shown some effects for vaccination and/or antigen-specific antibodies on several arthropod vectors control including sea lice [3, 4]. These results have encouraged the design of a cost effective process to produce the recombinant MY32/Cr for the development of a vaccine against sea lice.
The fermentation processes including a glucose batch phase followed by induction with isopropyl β-D-1-thiogalactopyranoside (IPTG) have been used to produce recombinant proteins in E. coli [5, 6]. However, they are time consuming and IPTG increases large scale production costs (can achieve up to 94 % of the culture medium cost). It has been previously demonstrated that the productivity of E. coli fermentation processes can be enhanced by using lactose as inductor by an auto-induction process [7].
On the other hand, an increase of serum IgM levels in teleost fish after immunization with several antigens has been referred [8, 9], but not always the IgM production has been protective. Nevertheless, some challenge experiments have demonstrated that the increase in IgM antibodies titers correlates with successful survival rates [10, 11].
Taking into account these facts, this work was aimed at improving production yields of the MY32/Cr recombinant protein, expressed in E. coli, by testing different fermentation culture media. The effect of the number of protein doses administrated on the IgM antibody response was also studied in Oreochromis niloticus as a model due to its advantages. IgM antibodies are produced by this other teleost fish as part of its acquired specific immunity, as in salmon. Moreover, O. niloticus is a fast growing fish, environmentally tolerant, disease and stress resistant, has a very low mortality, develops immune response relatively fast compared to salmon and has been used previously as animal model for different studies [12].
MATERIALS AND METHODS
Bacterial strain
The strain used in this study (BL21(DE3)-my32/Cr) was obtained from the transformation of the BL21(DE3) E. coli strain with the plasmid pET28a-my32 carrying the my32 gene from Caligus rogercresseyi [3].
Fermentation processes
Shake flasks containing 50 mL of medium inoculated with 50 µL from 20 % glycerol stocks of BL21(DE3)-my32/Cr [3] were used as inoculums for fermentation processes. Luria Bertani-Kanamycin (LBK) or non-inducible media were used to prepare the inoculums. The non-inducible medium was composed of: 20 mL of the 50×M medium per liter (177.5 g/L Na2HPO4, 170 g/L KH2PO4, 133.75 g/L NH4Cl, 80.5 g/L Na2SO4); 0.5 % (w/v) glucose; 0.25 % (w/v) L-aspartic acid; 0.492 g/L MgSO4×7H2O and 0.1 mM FeCl3-CaCl2.
Three medium compositions were tested for studying protein yield: 1) LBK; 2) complex medium I (CM I; 1 g/L NaCl/, 6 g/L NaH2PO4, 3 g/L K2HPO4, 1 g/L NH4Cl, 0.246 g/L MgSO4×7H2O, 0.01 g/L CaCl2, 0.01 g/L glucose, 10 g/L yeast extract, 100 µg/mL kanamycin) and 3) complex medium II (CM II), modified from Studier [7]: 20 mL of the 50×M medium per liter, 20 mL of the 5052 50× medium per liter (250 mL/L glycerol 98 %, 25 g/L glucose, 100 g/L α-lactose monohydrate), 0.492 g/L MgSO4×7H2O, 0.1 mM FeCl3-CaCl2, 10 g/L tryptone, 5 g/L yeast extract, 5 g/L NaCl, 100 µg/mL kanamycin. IPTG (1 mM; Promega, USA) was used to induce protein expression for LBK and CM I media. Lactose was used as inductor by an auto-inducing procedure for CM II medium.
Reagents employed to prepare culture media was supplied by AppliChem (Germany), except when specified. For LBK and CM I media, inoculums were grown on LBK medium. Cultures were grown until maximum specific growth rate as it was determined by a growth kinetics study performed previously; at that point, expression was induced by IPTG addition. Growth kinetics was checked by measuring optical density (600 nm) at different time points.
All cultures were performed in a 2-L BIOSTAT Plus bioreactor (Sartorius, Germany) containing 1 L of culture, to determine the influence of different culture conditions on the MY32/Cr yield, at 37 ºC and 600 rpm agitation. Aeration was set at 1 vvm. pH was adjusted to 7.0 ± 0.1 in all processes by the addition of 20 % (w/v) NaOH or 10 % (v/v) H3PO4; except when saline medium II was employed, in which pH was not controlled. Foam formation was avoided by adding Antifoam Glanapon DG 158 (Bussetti, Austria). After fermentation, the recombinant protein was purified as previously described [3].
Recombinant protein characterization by SDS-PAGE, Western blot and protein quantification
Protein expression was checked by SDS-PAGE (15 %) under reducing conditions and Western blot. The MY32/Cr protein was recognized with a specific MY32/Cr rabbit antiserum [3]. The blot was then developed with DAB (3,3′-diaminobenzidine; Sigma, USA) in the presence of H2O2 (Sigma, USA). For protein quantification, the MY32/Cr expression level (%) was determined by densitometry analysis from SDS-PAGE, using 1DManager program (TDI, Spain). Total protein concentration was determined by using the BCATM Protein Assay Kit (Pierce, USA).
Tilapia vaccination
Tilapias (Oreochromis niloticus) of average weight 80 g were distributed into three experimental groups (n = 5) and acclimatized in 400 L tanks one week prior to the experiment. Fishes were fed ad libitum with a basal diet (Cenpalab, Cuba) twice a day. The water temperature was maintained at 25-26 °C. Fish were immunized by intraperitoneal injection of 150 µL of the immunogen according to the experimental group. Fish in group I (control group) were immunized on day one with phosphate buffering saline solution (PBS 1×, pH 7.4). In groups II and III, fish were immunized on day one with 1 µg of the purified protein/g of body weight (gbw). On day 19 (500 degrees days) animals from group III received a booster with 1 µg/gbw. All vaccine preparations and the PBS 1× solution used for the control group were adjuvanted with Montanide 888 (50:50 v/v; SEPPIC, France). To detect specific IgM in serum, an ELISA was performed 28 days after the first immunization, using ELISA plates F96 (Nunc, Denmark). The plates were coated with 100 μL of purified antigen MY32/Cr (5 µg/mL) per well, formulated in sodium carbonate buffer (1.59 g/L Na2CO3, 2.93 g/L NaHCO3; pH 9.6). Serial dilutions of serum samples (100 μL/well; two replicates per dilution) were added to the plates, and reconstituted anti-O. niloticus IgMAb conjugated to horseradish peroxidase (C4-HRP, Aquatic Diagnostics, Ltd.) was further added according to manufacturer instructions for detection. Reactions were developed with 3,3’,5,5’-Tetramethylbenzidine (TMB). Endpoint titers were established as the reciprocal of the last dilution yielding an optical density (OD) higher than the mean OD of the pre-immune serum plus three times the standard deviation of pre-immune serum OD replica [13].
Statistical analysis
The statistical analyses were done using the GraphPad Prism statistical software, version 4.00.255 (GraphPad Software Inc., San Diego, California, USA). Previous to statistical analysis, a log2 transformation of the data was performed. They were further analyzed by one-way analysis of variance (ANOVA), followed by multiple comparisons by the Bonferroni test. Mean values were considered significantly different at p < 0.001.
RESULTS AND DISCUSSION
Yield improvement of MY32/Cr production in bioreactor
Recombinant MY32/Cr has been previously produced in E. coli using a standard IPTG induction protocol in 300 mL of LBK in shake flaks [3]. That study showed that the MY32/Cr protein yielded up to 50 mg/L on shake flasks, not feasible to develop a large scale fish vaccine production to meet the amount of market demands. Based on those results, improved conditions were established for culture in 1-L bioreactors, regarding medium composition and inductor.
Expression studies were performed using three culture media (LBK, CM I and CM II) employing IPTG or lactose as inductor. The expression kinetics shown by processes employing LBK o CM I culture media were the same observed for shake flask cultures (data not shown). The recombinant protein expression was detected 1 h after the addition of the inductor, and maximum expression was obtained 5 h after induction. When CM II (modified auto-induction medium containing glucose, glycerol and lactose [7]) was used, the expression of MY32/Cr was observed after 5 h since the beginning of the process, and maximum levels of expression were detected after 7 h (Figure 1A).

The identity of the expressed MY32/Cr protein was demonstrated by Western blot using a rabbit polyclonal antibody against MY32/Cr (Figure 1B). Cellular location studies, in fermentation and shake flask cultures, showed that the recombinant protein is produced intracellularly as inclusion bodies (data not shown).
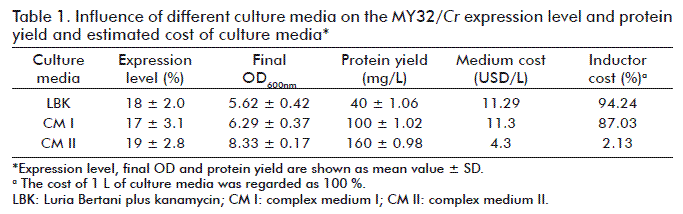
Culture medium composition did not affect protein expression levels, which was comparable in all cases. As shown in table 1, for all tested media MY32/Cr yielded approximately 20 % of the total cellular protein. It is advantageous that in the medium using lactose as inductor, the expression level was comparable to the media using IPTG as inductor, which is more expensive and can account for up to 94 % of the culture medium cost (Table 1).
The growth to high densities is desirable for maximizing the yield and efficiency of producing target proteins [7]. The OD values reached by the BL21 (DE3)-pET8a-my32/Cr strain were different in the tested media (Table 1, Figure 1C). As shown, CM II (complex medium containing glucose, glycerol and lactose which allows auto-induction) supported the highest final OD compared to LBK or CM I media, representing a notable influence of medium composition on final OD for the BL21(DE3) E. coli strain expressing MY32/Cr proteinn [14].
Similar results have been obtained by other authors for the expression of several proteins employing auto-induction media. It has been reported that auto-inducible cultures attain higher cell densities when saturated, in contrast to that using IPTG as inductor, for which final ODs are not usually higher than five initial OD units, and attaining high final cell densities when using this type of medium [15, 16]. Auto-induction is based on the function of the lac operon regulatory elements in mixtures of glucose, glycerol and lactose under diauxic growth conditions [17]. During the initial growth period, glucose is preferentially used as a carbon source and protein expression is low, due to catabolic repression of alternative carbon utilization pathways [18, 19] and binding interactions between lac repressors (LacI) and lac operators (lacO). As glucose is depleted, catabolite repression is relieved, which leads to a shift in cellular metabolism toward the import and consumption of lactose and glycerol. Lactose import results in the production of allolactose from lactose by a promiscuous reaction of β-galactosidase. Allolactose then acts as the physiological inducer of the lac operon. This supports the achievement of final high culture densities when auto-induction medium is employed, as achieved in this study, because of the advantage of the auto-induction method providing the transition from the un-induced to the induced state under metabolic control of the own host [7, 14].
Moreover, protein yields were dependent on culture medium composition and induction method. The highest was obtained with CM II medium, which was increased from that of CM I in more than 50 % (Table 1) and quadrupled that obtained with LBK medium, and also in shake flask culture. It has been reported the influence of medium composition not only on final OD, but also on protein yields in fermentation processes [15, 20]. The highest final OD values obtained with CM II made possible to obtain the highest biomass, and therefore, the highest yield, also considering that the protein is produced intracellularly. This in spite of the protein expression levels did not vary among the tested culture media. Increased yields of recombinant protein production, in the same order as MY32/Cr, have been recently reported with auto-induced medium for Arenicola marina globin chains (RecB2 and B2-GST), which were expressed at a yield of up to 105 mg/L of E. coli culture [21], for Bacillus halodurans β-glucosidase expressed in E. coli (468 mg/L) [22] and the green fluorescent protein (GFP), glutathione S-transferase-tagged GFP and other proteins (250 mg/L) [23].
The results obtained in this work showed that the use of auto-induction medium for expression provided the best results regarding OD and yield. It has been demonstrated that the auto-induction method has a good performance regarding cell mass recovered and yield at 37 °C for eGFP, luciferase and tobacco etch virus NIa proteinase [15, 16], and performed better than IPTG induction method [22].
Based on results obtained with the best performance medium in this study, and considering estimates of the annual world smolt salmon production, a tentative amount of MY32/Cr protein that should be produced annually for salmon vaccination against sea lice in aquaculture is presented in table 2. According to that data, up to approximately 150 kg of MY32/Cr protein are required to meet annual demands for this antigen for salmon vaccination.
Characterization of anti-MY32/Cr IgM immune responses in O. niloticus
Antibodies in the cutaneous mucus and skin of teleosts play a critical role in the protective immune response against infection. Fish are the most ancient bony vertebrates which contain immunoglobulins. However, they lack many isoforms which are present in mammals. Several Igs has been described recently in fish. Among them, IgM is the main Ig present in teleosts, appearing in different forms with a structural flexibility which is thought may compensate for a lack of diversity, shown to be functionally involved in protective immunity in fish [24, 25].
Considering this, the immunogenicity of the MY32/Cr protein, obtained in the improved fermentation processes established in this study, was determined by studying its ability to induce IgM immune responses in O. niloticus. Results showed that the intraperitoneal administration of MY32/Cr protein induced specific anti-MY32/Cr IgM antibodies (Figure 2).

Several authors have reported the significant increase of specific IgM titers after immunization for different antigens in Ictalurus punctatus, Salmo salar and O. niloticus [8, 11]. Group III, receiving a booster dose, showed IgM antibody titers higher than those of the group receiving a single dose (group II) and the control group (p < 0.001) (Figure 2). In this sense, Khushiramani et al. [9] showed that a booster dose of the ompTS antigen further increased the specific antibody production in immunization schemes, as compared to the response in animals receiving a single dose. Nevertheless, when these authors applied three booster doses, the fish showed lower titers, below those obtained in our work, which demonstrates that is not obvious that a single booster dose, as we performed, has to be highly immunogenic in all cases for this antigen. Furthermore, after 28 days, when several booster doses had been injected, IgM titers did not reach those attained by our team.
Moreover, in that study, no challenge experiment was reported supporting any correlation of booster application or antibody responses with challenge results. In our case, previous vaccination challenge trials performed in salmon with this recombinant protein using a single booster demonstrated to be successful (57 % of inhibition of sea lice infestation) [3]. Nevertheless, further immunological experiments in salmon are needed to corroborate this outcome. In fish, vaccines are normally administered early in the production cycle and, under ideal conditions, can provide protection throughout the growing cycle.
Revaccination strategies are also possible in marine finfish culture, and may provide a valuable method for reducing disease risk in long-lived, batch-spawning animals [26]. Re-vaccination is not a common practice for aquaculture vaccines for practical and economic reasons, but it is necessary when the primary immune response is weak and there is a high probability of re-infected fish. Also when the levels of immunity have decreased and the fish are environmentally challenged, such as during the seawater phase of salmon culture.
Noteworthy, there could be the possibility to provide a vaccine formulation able to increase IgM titers with a single re-immunization, as part of a set of factors to face sea lice infestations in the fish seawater growth phase. The results obtained in this work suggest that the increase of specific IgM titers when administering the MY32/Cr protein with a single booster could cooperate with fish protection against sea lice, but further experiments in salmon are needed to corroborate this outcome.
In summary, the expression level of MY32/Cr was not affected when it was expressed in different culture media, while using the CM II medium increased MY32/Cr yields up to four-fold those obtained in LBK medium in fermentation. One hundred and fifty kilograms was the estimated volume production of MY32/Cr protein to meet the annual demands for salmon vaccination. And the recombinant MY32/Cr obtained under the best performance fermentation conditions was highly immunogenic and induced IgM antibodies titers almost four-fold higher when administered twice (booster dose) than the single dose vaccination.
ACKNOWLEDGEMENTS
We would like to thank Raudel Sosa, Regla Margarita Somoza and Joana González for their useful and kind collaboration. This work was supported by a Research Project from Center for Genetic Engineering and Biotechnology.
REFERENCES
1. Costello MJ. The global economic cost of sea lice to the salmonid farming industry. J Fish Dis. 2009;32(1):115-8.
2. Goto A, Matsushita K, Gesellchen V, El Chamy L, Kuttenkeuler D, Takeuchi O, et al. Akirins are highly conserved nuclear proteins required for NF-kB dependent gene expression in drosophila and mice. Nat Immunol. 2008;9(1):97-104.
3. Carpio Y, Basabe L, Acosta J, Rodríguez A, Mendoza A, Lispergerb A, et al. Novel gene isolated from Caligus rogercresseyi: A promising target for vaccine development against sea lice. Vaccine. 2011;29(15): 2810-20.
4. Manzano-Román R, Díaz-Martín V, Oleaga A, Siles-Lucas M, Pérez-Sánchez R. Subolesin/akirin orthologs from Ornithodoros spp. soft ticks: cloning, RNAi gene silencing and protective effect of the recombinant proteins. Vet Parasitol. 2012;185(2-4):248-59.
5. Volontè F, Piubelli L, Pollegioni L. Optimizing HIV-1 protease production in Escherichia coli as fusion protein. Microb Cell Fact. 2011;10:53.
6. Hortsch R, Weuster-Botz D. Growth and recombinant protein expression with Escherichia coli in different batch cultivation media. Appl Microbiol Biotechnol. 2011;90(1):69-76.
7. Studier FW. Protein production by auto-induction in high-density shaking cultures. Prot Expr Purif. 2005;41(1):207-34.
8. Wilhelm V, Soza C, Martínez R, Rosemblatt M, Burzio LO, Valenzuela PD. Production and immune response of recombinant Hsp60 and Hsp70 from the salmon pathogen Piscirickettsia salmonis. Biol Res. 2005;38:69-82.
9. Khushiramani R, Girisha SK, Karunasagar I. Cloning and expression of an outer membrane protein ompTS of Aeromonas hydrophila and study of immunogenicity in fish. Protein Exp Purif. 2007;51(2):303-7.
10. Wang X, Dickerson HW. Surface immobilization antigen of the parasitic ciliate Ichthyophthirius multifiliis elicits protective immunity in channel catfish (Ictalurus punctatus). Clin Diagnos Lab Immunol. 2002;9(1):176-81.
11. Xu DH, Klesius PH, Shoemaker CA. Protective immunity of Nile tilapia against Ichthyophthirius multifiliis post-immunization with live theronts and sonicated trophonts. Fish Shellfish Immunol. 2008;25(1-2):124-7.
12. Chuang WL, Sun B. Anti-stress effects of Glycine tomentella Hayata in tilapia: Inhibiting COX-2 expression and enhancing EPA synthesis in erythrocyte membrane and fish growth. Agric Food Chem. 2011;59(17):9532-41.
13. Ochoa RF. Guide for immunoenzimatic technique standarization for vaccine trials. In: Ochoa RF. Methodological bases for antibody evaluation in vaccine clinical trials by immunoenzimatic techniques. 2nd Ed. La Habana: Finlay Ediciones; 2008. p. 31-45.
14. Sreenath HK, Bingman CA, Buchan BW et al. Protocols for production of selenomethionine-labeled proteins in 2-L polyethylene terephthalate bottles using auto-induction medium. Protein Exp Purif. 2005;40(2):256-67.
15. Blommel PG, Becker KJ, Duvnjak P, Fox BG. Enhanced bacterial protein expression during auto-induction obtained by alteration of Lac repressor dosage and medium composition. Biotechnol Prog. 2007;23(3):585-98.
16. Blommel PG, Fox BG. A combined approach to improving large-scale production of tobacco etch virus protease. Protein Exp Purif. 2007;55(1):53-68.
17. Jacob F, Monod J. Genetic regulatory mechanisms in the synthesis of proteins. J Mol Biol. 1961;3:318-56.
18. Inada T, Kimata K, Aiba H. Mechanism responsible for glucose-lactose diauxie in Escherichia coli: challenge to the cAMP model. Genes Cells. 1996;1(3):293-301.
19. Hogema BM, Arents JC, Bader R, Postma PW. Autoregulation of Lactose uptake through the lacY permease by enzyme IIAGlc of the PTS in Escherichia coli K-12. Mol Microbiol. 1999;31(6):1825-33.
20. Berger C, Montag C, Berndt S, Huster D. Optimization of Escherichia coli cultivation methods for high yield neuropeptide Y receptor type 2 production. Protein Expr Purif. 2011;76(1):25-35.
21. Harnois T, Rousselot M, Rogniaux H, Zal F. High-level Production of recombinant Arenicola marina globin chains in Escherichia coli: A new generation of blood substitute. Artif Cells Blood Substit Immobil Biotechnol. 2009; 37(3):106-16.
22. Naz S, Ikram N, Rajoka MI, Sadaf S, Akhtar MW. Enhanced production and characterization of a beta-glucosidase from Bacillus halodurans expressed in Escherichia coli. Biochemistry (Mosc). 2010;75(4):513-25.
23. Li Z, Kessler W, van den Heuvel J, Rinas U. Simple defined autoinduction medium for high-level recombinant protein production using T7-based Escherichia coli expression systems. Appl Microbiol Biotechnol. 2011;91(4):1203-13.
24. Solem ST, Stenvik J. Antibody repertoire development in teleosts–a review with emphasis on salmonids and Gadus morhua L. Dev Comp Immunol. 2006;30(1-2):57-76.
25. Zapata A, Amemiya CT. Phylogeny of lower vertebrates and their immunological structures. Curr Top Microbiol Immunol. 2000; 248:67-107.
26. Bricknell IR, Bron JE, Bowden TJ. Diseases of gadoid fish in cultivation: a review. J Mar Sci. 2006;63(2):253-66.
Received in March, 2013.
Accepted in June, 2013.
Mario P Estrada. Departamento de Biotecnología Animal, Dirección de Investigaciones Agropecuarias, Centro de Ingeniería Genética y Biotecnología, CIGB. Ave. 31 e/ 158 y 190, Cubanacán, Playa, CP 11600, La Habana, Cuba. E-mail: mario.pablo@cigb.edu.cu.

{kind=link}
{kind=link}