










My SciELO
 Custom services
Custom servicesServices on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkBiotecnología Aplicada
On-line version ISSN 1027-2852
Biotecnol Apl vol.31 no.2 La Habana Apr.-June 2014
REVIEW
MicroRNAs as biomarkers and therapeutic targets in cancer
Los microARN como biomarcadores y blancos terapéuticos en cáncer
Alejandro Moro-Soria
Instituto Nacional de Oncología y Radiobiología, INOR. Calle 29 y F, CP 10400, Plaza de la Revolución, La Habana, Cuba.
ABSTRACT
Malignant tumors rank as the first cause of death mortality and potentially lost years of life in Cuba and many other countries where millions of people die each year because of cancer. Therefore, the diagnosis and treatment of cancer is a health issue worldwide. Malignant tumors are traditionally classified and treated according to clinical and pathological variables which are currently insufficient to reflect each patient’s molecular features. Consequently, there is a need to find new individual biomarkers and therapeutic targets that could complement current clinical-pathological variables and become a guide for diagnosis, prognosis and treatment. Advances in understanding the biology of cancer as well as the development of more powerful gene analysis tools create new clinical perspectives to find such biomarkers and targets. MicroRNAs (miRNAs) belong to a class of non-coding RNA with post-transcription regulation of gene expression. Their expression is aberrant in cancer where they could function either as oncogenes or tumor suppressors, being used as diagnostic, prognostic or predictive biomarkers and therapeutic targets. Since, research on miRNA is a key issue for oncologists and researchers, in this review we provide a description of their biogenesis, mechanism of action and recent findings, supporting their use in cancer, either as biomarkers or therapeutic targets.
Keywords: miRNA, biomarkers, cancer, therapeutic molecular targets, personalized oncology.
RESUMEN
Los tumores malignos son la primera causa de muerte temprana de millones de personas en Cuba y otros países, por lo que el diagnóstico y tratamiento del cáncer es un problema de salud mundial. Tradicionalmente, estos tumores se clasifican y tratan según variables clínico-patológicas un tanto imprecisas todavía, pues no reflejan las particularidades moleculares de cada paciente. En este sentido, es preciso encontrar otros biomarcadores individuales y blancos terapéuticos que complementen esas variables y guíen el diagnóstico de los pacientes, su pronóstico y tratamiento. Los avances en el conocimiento de la biología del cáncer y el desarrollo de herramientas para los análisis genómicos, constituyen perspectivas clínicas para el descubrimiento de estos biomarcadores. Los microARN (miARN) son un tipo de ARN no codificante que provocan la regulación postranscripcional de la expresión génica. Su expresión en el cáncer está alterada, y podrían funcionar como oncogenes o supresores de tumores, por lo que se utilizan como biomarcadores diagnósticos, pronósticos, predictivos y blancos terapéuticos. Teniendo en cuenta la importancia de este tema para oncólogos e investigadores, se describen la biogénesis de los miARN, su mecanismo de acción y los avances en el estudio como biomarcadores o dianas terapéuticas en el cáncer.
Palabras clave: miARN, biomarcadores, cáncer, blancos moleculares terapéuticos, oncología personalizada.
INTRODUCTION
The ideal biomarker must be specific for the disease of interest, available by non-invasive techniques and detected by methods that are not too costly. Those biomarkers, which may distinguish the molecular bases of variations in tumor evolution and the clinical response to treatments, are of great medical value [1]. According to the database of GLOBOCAN, from the International Agency for Research on Cancer (IARC), cancer is a growing health problem; its incidence has increased from 12.7 million in 2008 to 14.1 million in 2012, and mortality has increased from 7.6 million to 8.2 million [2]. In 2012, malignant tumors in Cuba were the first cause of death (22 532 deaths), exceeding those due to cardiovascular diseases (22 234 deaths), and the first cause of potentially lost years of life (18.2 years) [3]. This shows the importance of finding new biomarkers for the early detection and characterization of tumors, as in the use of the RNA molecules for this purpose.
In the last twenty years, new types of RNA have been characterized through their gene expression regulating functions and variable size. These include: 1) siRNA (small interference RNA) of 20-25 nucleotides, related to the defense of the body against viral infections; 2) RNA associated to Piwi (piRNA) of 25-30 nucleotides, that operates in association with the Piwi subfamily of Argonaute proteins, and are essential for the development of germ cells; 3) microRNA (miRNA) of 20 to 25 nucleotides that are related to the repression or degradation of the messenger RNA (mRNA) in several species; and 4) long non-coding RNA (LncRNA), of variable size, from 70 to several thousand nucleotides, which participate in several cellular processes, including maturation of the mRNA and the biogenesis of the ribosomes [4]. The miRNAs are of particular interest due to their relevance in cancer studies.
The control of genetic expression by miRNAs occurs in all tumor cells. These cells frequently have an alteration of the expression profile of miRNAs, and recent evidence indicates that these alterations may have diagnostic, prognostic and predictive value for cancer [5]. In recent years, there have been advancements in the progression mechanisms and development of therapeutic strategies to prevent the dissemination of cancer throughout the body. This made it possible to increase the percentage of patients with a 5 year survival after diagnostics. However, metastasis is the main cause of death of these patients. It is therefore essential to identify metastasis mediators, which can be used as biomarkers or therapeutic targets. This information would be a useful tool for better clinical decision making in cancer, thus adjusting the treatments to the molecular specificities of each patient. In this context, the miRNAs represent a new class of potential biomarkers and therapeutic targets.
In this paper, we give examples of the functions of the miRNAs as possible biomarkers for the diagnostic of lung cancer, the prognosis of breast cancer, and as predictive biomarkers in colon cancer, since these three locations are the main causes of death worldwide and in Cuba. Their potential as therapeutic targets is also shown, including their advantages as circulating biomarkers, and the limitations of their use. Finally, we offer an analysis of their current situation and the perspectives of working with them.
DISCOVERY AND GENOMIC STRUCTURE
The first miRNAs discovered were the lin-4 (derived from abnormal lineage) and let-7 (derived from lethal), in the worm Caenorhabditis elegans, led by Ambros and Ruvkun‟s groups in 1993 and in 2000, respectively [6, 7]. They found that these were atypical RNA, because they were produced as a strand of 22 (lin-4) or 21 (let-7) nucleotides (nt); they were derived from precursors of 70 nucleotides (nt) with the stem-loop structure, and their functions were of antisense repressors of mRNA translation, coding for the proteins needed for the development of C. elegans in the correct sequence.
When they were described, lin-4 and let-7 seemed to be exclusive for C. elegans, a rareness, since no other small RNA had been described with a regulating function in any species. Two discoveries, however, changed this perception. The first one was that the RNA of let-7 is phylogenetically preserved, in size and sequence, in all bilaterally symmetric animals [8]; and the second finding was that the small size antisense RNA, of approximately 22 nt (called siRNA) were essential for the RNA interference (RNAi) process [9].
RNAi is a genetically preserved surveillance mechanism that may degrade the mRNA, as a response to the presence of double strand RNA, corresponding to the target mRNA. The lin-4 and let-7 are not siRNA, since their action does not need the degradation of their target mRNA; but the ubiquity of the siRNA suggested that small size RNAs such as lin-4 and let-7 are quite stable in the eukaryotic environment, in which they remain for a relatively long period. In fact, the lin-4 and let-7 RNA, of approximately 22 nt, are processed by means of their precursors, in the stem-loop form, by the same Dicer enzyme generating the siRNA with a size of approximately 22 nt from double strand RNA, a mechanism that is explained later on. Considering that the Dicer enzyme is also widely distributed, it was presumed that molecules similar to lin-4 and let-7 may be found in other organisms besides C. elegans. This hypothesis was confirmed in 2001, which led to three papers published in the same issue of Science, describing more than 100 miRNAs in several metazoan cells [10-12]. These papers had a strong impact, since a new path in genetic regulation was opened on incorporating miRNA.
It is now known that the eukaryotic cells have siRNA or at least the cellular machinery necessary to produce them, with the exception of the Saccharomyces cerevisiae yeast. The miRNAs basically operate in multicellular organisms, since they were also identified in the unicellular algae Chlamydomonas reinhardtii [13]. And although it was not so surprising, in 2004, it was confirmed that some viruses contained miRNAs [14]. This demonstrated that they could be of great importance for the regulation of the transition from the latency to the replication and lysis stages; and they can even act in the attenuation of the immune response of the host against viral infection.
Most of the miRNAs characterized are located in the intergenic space, or in the opposite direction to neighboring genes. Therefore, it was thought that they were transcribed as independent units [15]. However, in some cases the miRNA is transcribed together with the neighboring gene. This indicates that the expression of the miRNA and of the protein-coding neighboring genes may be associated [16].
The large scale genomic analyses of the miRNAs show that most of them form clusters or groups of genes; they are polycistronic and are coexpressed with neighboring miRNAs [17]. It was also demonstrated that these clusters of miRNA cooperate and coordinate their functions, thereby being more efficient in the regulation of genes [18, 19] and their related proteins [20]. The hypothesis is that the grouped miRNAs are functionally related, because their target is the same gene or perhaps several genes, but
they encode related proteins [21]. There is evidence supporting this hypothesis. For example, for a cell to proliferate and invade other tissues, it has to avoid the control mechanisms such as cellular arrest and apoptosis. It was demonstrated that miRNAs 15a and 16, which form a cluster in chromosome region 13q14 (frequently eliminated in several neoplasia), act as suppressors of prostate cancer tumors, because they have the bcl-2 genes, of the cyclin D1 and wnt3A as their target. These genes codify proteins that inhibit apoptosis, promote cellular proliferation and invasion, respectively. In advanced prostate cancer the concentrations of miR15a and miR16-1 are very low, while those of Bcl-2, cyclin D1 and Wnt3A are high [22].
NOMENCLATURE
The adoption of a system to identify miRNAs facilitated their study, together with the implementation of a miRNA database or registry [23]. The miRNA names are assigned following experts guidelines, which state that new miRNAs must be experimentally verified by cloning or through expression and function evidences [24]. The name starts with the mir- prefix followed by the number of its sequential identification. For instance, mir-100 preceded mir-101. The prefix mir- identifying the precursor miRNA is changed to miR (capital letter) to indicate the mature form of the molecule. When two miRNAs are almost-identical but differing in just one or two nucleotides, a lower case letter is added after the number, i.e., miR-123a is almost identical to miR-123b.
Moreover, those miRNAs located in different genomic regions but producing an identical mature miRNA are denoted by an additional hyphen at the end, followed by a sequential number. For example, mir-194-1 and mir-194-2 produce identical mature mRNAs, miR-194. Their genes are in different genome locations, though. The species are further indicated by a three-letter prefix, i.e., the hsa-miR-100 is the one-hundredth miRNA isolated in humans (Homo sapiens), while the mmu-miR-100 is the one-hundredth identified in mouse (Mus musculus).
The miRNA numbering is sequential, so, if the last discovered miRNA was miR-789 in mice, the next name published will be miR-790 regardless the species where it was discovered. If a sequence is submitted for a new miRNA identified in Gallus gallus but identical to that of the human miR-121, it will be named miR-121, with the prefix gga-121 different from the has-121 one.
If two different miRNAs are originated in opposite arms of the same precursor (or pre-mir), they will be denoted by the suffixes -3p or -5p. In those cases of known expression levels, an asterisk is added at the end of the name of the miRNA displaying the lowest expression level, compared to that in the other arm of the same precursor. For example, miR-234 and miR-234* share the same precursor, mir-234; but it is found at higher concentrations within the cell.
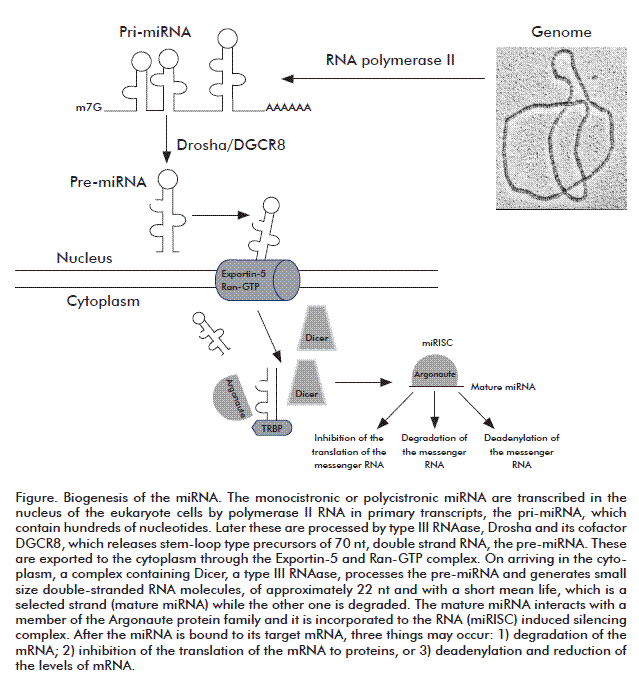
BIOGENESIS
The general mechanism of biogenesis of the miRNA is described in the figure. These are transcribed as part of the major primary transcripts (pri-miRNA), generated by the action of RNA polymerase II [25]. As in the other RNA polymerase II transcripts, the pri-miRNAs have their 5´ and 3´ ends modified with CAP and a polyA tail, respectively [26]. These pri-miRNAs form a stem-loop structure recognized by a multiprotein system with two main components, the Drosha enzyme and the DGCR8 protein (Di George syndrome critical region gene 8), capable of binding to double-stranded RNA. In this recognizing mechanism, DCGR8 is bound to the base of the stem-loop structure, thereby guiding the Drosha enzyme that is a type III RNAase, which has the catalytic activity of the complex [27]. Drosha cuts the double strand at the stem, at an approximate distance of 11 nt above the base and generates 3´ ends with overhangs of 2 nt. This cut releases an RNA molecule, with the stem-loop form, of approximately 70 to 100 nt, which constitutes the precursor (pre-miRNA) and it is recognized by the nuclear receptor Exportin-5, and is exported from the nucleus to the cytoplasm [28]. After its arrival at the cytosol, the pre-miRNA is again processed by a type III RNAase, Dicer [29]. The PAZ dominion of this enzyme is bound to the 3´ end with the 2 nt overhang of the pre-miRNA (substrate), by which means the substrate is in the appropriate position for cutting, resulting in a double-stranded miRNA of 22 nt with 2 ntoverhangs at the 3´ ends [30]. One of the strands, the leading strand or mature miRNA, is transferred to a protein named Argonaute and a complex is formed called miRISC (miRNA induced silencing complex), while the other one is degraded. In mammals, the complex formed contains mature miRNA, the Dicer enzyme, the Argonaute protein and another protein that binds to double stranded RNA, TRBP [31]. In general, the miRNA strand, having less stability in base-pairing at its 5´ end, is the guide strand or mature miRNA [32].
MECHANISM OF ACTION
After the formation of the RISC complex, it is bound to the 3´ untranslated region (3´-UTR) of the mRNA, although it can also be bound to the 5´ untranslated region (5´-UTR) and the promoter region. The sequence in the miRNA that recognizes the 3´-UTR of the mRNA is known as the “seed region” and it is located at the 5´ region of the miRNA, of nucleotide 2 to 7 [33]. After the miRNA is bound to its mRNA target, there may be a degradation of the mRNA, or the inhibition of the translation of mRNA to proteins, or the deadenylation and reduction of the levels of mRNA [34].
THE miRNAs AS BIOMARKERS IN CANCER
The early diagnosis of a primary tumor improves the prognosis and response to treatment. Since there is no system that can be used to screen lung cancer in its early stages within the population, it is mostly diagnosed at advanced stages. The first evidence of an aberrant expression of the miRNAs in lung cancer was reported in 2006, when a group of them was identified, which were expressed differently in the tumor tissue compared to healthy tissues [35]. This opened the possibility of their use as biomarkers of this type of cancer. Recent studies demonstrated that the increase in the expression of the miR-21 was significantly different in the sputum of patients with lung cancer compared to healthy persons, with a sensitivity of 70 % and a specificity of 100 % [36]. In experiments with rats it was also found that the exposure to cigarette smoke modified the levels of expression of the miRNA [37], indicating that the changes in the levels of the miRNA can be used as biomarkers for cancer diagnosis in smokers.
The ten miRNAs with aberrant expressions in tumor tissues, which can be used to confirm the diagnostic of lung cancer through computerized tomography, and very valuable in discriminating neoplastic from healthy tissues are: let-7, miR- 21, -200b, -210, -219-1, -324, which are (induced); and miR-30a, -126, -45 and -486, which are (repressed) [38]. To develop therapies for specific targets, tumors must be classified as: non-small-cell lung cancers (NSCLC), the adenocarcinoma histological subtypes (ADC) and the squamous cell carcinomas (SCC). This, however, is not easy to do, especially if the biopsy is small or the tumors are hardly differentiated. In this sense, two miRNA, miR-21 and -205 were successfully used to distinguish ADC from SCC [39]. A characteristic expression pattern of five miRNA (miR- 25, -34c-5p, -191, let-7e and -34a) used for this differentiation was also described [40]. It is also necessary to distinguish the primary tumors from the metastases. The overexpression of miR-182 was detected in primary tumors, while miR-126 was expressed in high concentrations in the metastases [41].
The main cause of death of patients with breast cancer is due to the complications derived from the metastases [42]. In recent years, a group of studies were carried out to identify and characterize the very complex cellular events that rule the process of metastasis. The miRNAs affect cellular adherence, migration, invasion, motility and angiogenesis [43]. Those associated to the metastasis process are called metastamirs. The first metastamir was described by Robert Weinberg and his group, who found that miR-10b starts the invasion and the metastasis process in breast cancer [44]. The first metastasis suppressor metastamir was described by Joan Massagué and his group [45], revealing that miR-335 suppresses metastasis and migration through its action on the SOX4 transcriptional factor, found in stem cells and on the tenascin C protein, which forms part of the extracellular matrix.
Metastamirs are used in breast cancer as prognostic biomarkers. For example, in a microarray study of miRNA, the expression of 249 miRNA was quantified in 299 breast tumors (185 were positive to the estrogen receptor, RE+; 114 negative, RE-). The miRNAs -7, -128a, -210 and -516-3p were related to the aggressiveness of the tumor in RE+ tumors and negative nodules, while miR-210 was associated to an early relapse in patients with RE- and negative nodules [46]. In breast cancer it was observed that miRNA 27 and those of the 17-92 family (miR-17, -18a, -19a, -19b, -20a and -92) negatively regulate the concentrations of the transcriptional factors of the ZBTB family, and that these concentrations are significantly associated to survival [47, 48].
The survival of 466 patients with invasive ductal carcinoma (IDC), the most frequently diagnosed histological subtype, was recently carried out through the integration of the expressions of mRNA, miRNA and methylated DNA [49]. In the analysis of this integrated genetic pattern, it was found that 37 mRNA and 7 miRNA, within the clinical and pathological subclasses, are associated to general survival and that their prognostic value is higher than other commercially available genetic patterns for the stratification of patients with breast cancer, such as Mammaprint® and Oncotype Dx®.
The progress in the knowledge of the altered signaling routes in colorectal cancer (CRC), laid the foundations for the application of therapies with specific targets. For example the cetuximab monoclonal antibody against the epidermal growth factor receptor (EGFR) is used as a monotherapy against the CRC [50], or it is combined with chemotherapy [51]. It was recently known that patients with mutations in the K-ras gene (approximately 38 %) are resistant to the anti-EGFR therapy [52]. This therapy is therefore restricted to the subgroup of patients with CRC not having the K-ras gene mutation, although this is not an effective treatment for all of them and it would be necessary to find new markers. It was recently described that the miRNA let-7 [53], miR-143 [54] and miR-18a [55], with reduced concentrations in CRC, have K-ras as one of their target genes. In this sense, the predictive ability of 18 miRNA (several of which have the K-ras as their target) in 32 patients with metastatic CRC (CRCm) treated with cetuximab and chemotherapy, was evaluated. The increase in the expression of miR-200 and the decrease in that of miR-143, were associated with a longer period of time in which the patients with the mutated K-ras gene were free from progression. There was no correlation between the expression of the miRNA with the K-ras gene as the target, and the time that the patients with non-mutated K-ras were free from progression. Therefore, the inhibition of the expression of the K-ras gene by the miRNA is only relevant in the context where there are high levels of K-ras; i.e., when there are activator mutations [56].
In a study of the expression profiles of miRNAs in 99 patients with CRCm, 33 of them with non-mutated K-ras/BRAF genes, to predict those having a favorable response with cetuximab and a longer survival, it was found that patients with the progressive disease had an overexpression of miR-31 and an inhibition of miR-592. Patients were also grouped on the basis of the miRNAs expression profile and significant differences between groups were found. The patients with short survival had the miRNAs of the let-7 family, the miR-140-5p overexpressed and the miR-1224-5p inhibited [57].
The standard treatment for those with locally advanced CRC (approximately one third of all patients with CRC), are the neoadjuvant radiochemotherapies with capecitabine or 5-fluorouracil, and later surgical resection. Unfortunately, a large percentage of these tumors do not respond to the neoadjuvants and patients have high probabilities of having tumor relapses.
In a recent study, the profiles of the miRNAs were analyzed in 20 patients with tumors classified as sensitive or resistant to that therapeutic variant. The levels of expression of eight miRNAs were significantly different between both groups. The miRNAs -215, -190b and -29b2* were overexpressed in patients with resistant tumors, while the let-7e, miR-196b, -450a, -450b-5p and -99a* were overexpressed in sensitive tumors. The levels of expression of these miRNA enabled the classification of nine patients (out of 10) as non-responders, and the same number as responders. It was concluded that the miRNA form part of the response mechanism of the cancer cells of the rectum to radiochemotherapy, and therefore, they are considered to have an important potential as predictive markers in patients diagnosed with this neoplasia [58].
THE miRNAs AS THERAPEUTIC TARGETS IN CANCER
The miRNAs have two characteristics that make them attractive as therapeutic targets for cancer. One of them is that they regulate several metabolic pathways: the complexity and heterogeneity of cancer suggests that the only way to develop a successful treatment is considering multiple genes simultaneously as targets. The second one is that there are microRNAs that are aberrantly overexpressed in cancer and they are known as oncomirs. Therefore, it may be easier and economically more attractive to develop drugs against the miRNA, which would be potentially useful to treat malignant tumors in several localizations. The inhibition of the miRNA may be achieved by antisense oligonucleotides or antagomir [59, 60] or competitive inhibitors called miRNA “sponges” [61]. These contain several binding sites to the miRNA, whether in a non-codified transcript or in the 3´-UTR of a reporter gene, and they have advantages in relation to the antagomir.
First of all, the antagomirs depend, for their effectiveness, on an extensive complementarity zone with the miRNA target [62]. Therefore, to neutralize several miRNAs a mixture containing antagomirs that are complementary to these miRNAs must be prepared, while if an mRNA containing multiple binding sites (typically 4 to 10 sites) to the miRNA of interest is introduced into the cell, it will specifically inhibit all the miRNAs having a complementary sequence to these binding sites.
In the second place, many cells are resistant to the introduction of the antagomirs, while the genetic construction containing the sponge can be introduced in the cell through several types of vectors. Thirdly, the antagomirs must be periodically administered at high concentrations to inhibit the expression of the target miRNA; while the expression of the genetic construct of the sponge in germ cells of transgenic animals would make it possible for the miRNA of interest to be inhibited for the entire life span of the animal [63]. However, although the inhibition technology of the miRNA sponge has advantages in experimentation with biological models, the inhibition using antagomirs is considered to be more attractive and has a greater potential from the therapeutic viewpoint. In fact, the first molecule targeting a miRNA in human beings, for the treatment of hepatitis produced by the hepatitis C virus (HCV), was Miravirsen [64] (originally called SPC3649 [65]; Santaris Pharma A/S). It is an antagomir against miR-122, a positive regulator of the replication of HCV and a tumor suppressor in liver carcinoma.
In 2005, it was demonstrated that the miR-122 has two seed sites, which interact with the 5´-UTR region of the RNA of the HCV, and produce the increase of viral RNA in infected liver cells. This evidence indicates that the miR-122 is an important factor in the host and a potential target for the treatment of infections due to the HCV; this is relevant considering there are approximately 170 million persons in the world infected with HCV and it is the main cause of liver failure [66]. The idea of developing an antiviral therapy, based on miR-122 as the target, was attractive because of the limitations of the current treatments for the infections due to HCV. Besides its importance as the first anti-miRNA drug, there are another two attractive characteristics of Miravirsen. This antagomir was used to demonstrate that on modulating a miRNA with this type of molecule, the genes downstream the miRNA are also regulated. Furthermore, Miravirsen is a molecule with a chemical structure having a modification to increase its stability, affinity and specificity. Such modification consists of closing the ribose ring through a methylene bridge, which connects the oxygen at position 2 with the carbon at position 4, producing a locked nucleic acid (LNA). The LNAs are nucleotides with a high hybridization capacity and affinity for the single stranded RNA [67]. Therefore, the miRNAs modified with LNA, such as Miravirsen, offer encouraging perspectives for clinical practice.
At present, the biotechnology companies are developing miRNA or antagomir mimetics, and strategies for their systemic administration in cancer treatment and other diseases [64]. Taking into account the strong relationship between the miRNA and the deregulation of the malignant processes, a faster approval for the use of therapeutic strategies based on its modulation in monotherapies or combined with radio-, chemo-, or immunotherapies, are foreseen.
CIRCULATING miRNAs
The description of the circulating miRNA meant the discovery of new mediators of the genetic expression. Although most of them are found and function inside the cell, the circulating miRNAs can also be found outside cells: in fluids such as plasma, saliva, amniotic fluid, urine and others [68]. Their composition and concentrations are related to several diseases, and they can therefore be used as biomarkers, measured through pathways that are hardly invasive, to assess and keep watch over the physiological state of the body. It is important to highlight that these miRNAs may be found in large amounts in extracellular fluids, and may be very stable, even under conditions of extreme temperature, pH and storage. Their composition in the plasma is identical in all persons, not depending on sex or age, so they are considered to be good clinical biomarkers [69]. It is not well defined, however, how their action is exerted. It has been said that they could be signal mediators, allowing the tumor cells to „communicate‟ with the non-transformed cells from the microenvironment, which is crucial in malignant progression. Therefore, the molecules that transport any type of information to the microenvironment, such as cytokines, chemokines and growth factors, are bound to circulating miRNAs [70].
LIMITATIONS IN THE USE OF miRNAs AS BIOMARKERS
The introduction of new biomarkers in clinical practice is limited because of insufficient sensitivity, specificity and stability of the potential markers.
Although miRNAs are considered biomarkers with a great potential, they also have certain limitations, such as their specificity. The miR-21 is high, not only in patients with pancreas cancer, but also in those with diffuse large B-cell lymphoma [71]. Another important aspect is how to obtain it, since no homogeneity is found in the protocols for their isolation and characterization, the starting material (whether it is plasma, serum, tissue or paraffin blocks), and the characteristics of the patients included in the studies. There is evidence that erythrocytes are an important source of circulating miRNAs and that hemolysis can alter up to 50 % the levels of a certain miRNA, as a biomarker in the plasma. Therefore, one must be very rigorous when interpreting the results of the mediation of the circulating miRNA associated with cancer, to figure out if they are really related to the malignant process and not to an event that is a consequence of cellular lysis. Another aspect that must be standardized is the quantification of the miRNAs. There is no consensus on which molecule to use as the gene for normalization: whether it is a miRNA that does not vary, another RNA molecule or another gene.
FINAL COMMENTS
In the last two decades, there has been a significant progress in the identification and characterization of genetic and epigenetic causes of cancer, in the assessment of possible biomarkers, and in the development of drugs against new targets, which had an impact on the relatively insufficient increase of survival time of oncological patients after their diagnosis. A greater in-depth knowledge of cancer biology may help identify other alterations that, by causing or contributing to malignancy, may be used as new biomarkers or molecular targets and continue increasing survival time of the patients. In this sense, it was recently found that the miRNAs are essential molecules with important functions in the control of several physiological and pathological processes (including cancer), with a differentially modulated expression in healthy tissues or in those undergoing a pathological process not associated to malignancy, and correlated to clinical-pathological characteristics.
The high stability of the miRNAs in tissues and body fluids, and the possibility of being quantified, although they are found in small amounts, are advantages for diagnosis, prognosis, and stratification of patients, with the aim of increasing the objective responses to antineoplastic treatments. For example, the miRInform® Pancreas LDT (Asuragen® Inc) system [72] is already on the market; it was developed after the analysis of the differential expression of the miR-196a and the miR-217 [73, 74]. Through a biopsy, this system makes it possible to discriminate chronic pancreatitis from pancreatic adenocarcinoma. Furthermore, the ability of each miRNA to modulate the expression of multiple mRNA at the same time is also attractive, since it enables, for example, to re-establish functionality of a complete suppressor route that had been affected, in contrast to other strategies that would only allow to modulate the expression of a gene but not involve the entire route. This feature makes them very attractive as therapeutic targets themselves, or to decrease the resistance of other treatments; these are aspects under intense study by academic researchers or the industry.
In the next 2 to 5 years an increase in the systems based on miRNAs approved by the regulating agencies is foreseen. They may be introduced in routine clinical practice to increase the sensitivity and specificity of the diagnostic and prognostic tests, which may offer new opportunities for better personalized treatments.
REFERENCES
1. Biomarkers Definitions Working Group. Biomarkers and surrogate endpoints: preferred definitions and conceptual framework. Clinical Pharmacol Ther. 2001;69(3):89-95.
2. Ferlay J, Soerjomataram I, Ervik M, Dikshit R, Eser S, Mathers C, et al. GLOBOCAN 2012 v1.0, Cancer Incidence and Mortality Worldwide: IARC CancerBase No. 11 [Internet]. Lyon, France: International Agency for Research on Cancer. c2013 [cited 2013 Oct 17]. Available from: http://globocan.iarc.fr.
3. Ministerio de Salud Pública. Anuario Estadístico de Salud, Cuba, 2012 [Internet]. Habana: Dirección Nacional de Registros Médicos y Estadísticas de Salud, Minsap; 2013 [cited 2013 Oct 17]. Available from: http://files.sld.cu/dne/files/2013/04/anuario_2012.pdf.
4. Wang J, Li LC. Small RNA and its application in andrology and urology. Transl Androl Urol. 2012;1(1):33-43.
5. Lujambio A, Lowe SW. The microcosmos of cancer. Nature. 2012;482(7385):347-55.
6. Lee RC, Feinbaum RL, Ambros V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell. 1993;75(5):843-54.
7. Reinhart BJ, Slack FJ, Basson M, Pasquinelli AE, Bettinger JC, Rougvie AE, et al. The 21-nucleotide let-7 RNA regulates developmental timing in Caenorhabditis elegans. Nature. 2000;403(6772):901-6.
8. Pasquinelli AE, Reinhart BJ, Slack F, Martindale MQ, Kuroda MI, Maller B, et al. Conservation of the sequence and temporal expression of let-7 heterochronic regulatory RNA. Nature. 2000;408(6808):86-9.
9. Sharp PA. RNA interference--2001. Genes Dev. 2001;15(5):485-90.
10. Lagos-Quintana M, Rauhut R, Lendeckel W, Tuschl T. Identification of novel genes coding for small expressed RNAs. Science. 2001;294(5543):853-8.
11. Lau NC, Lim LP, Weinstein EG, Bartel DP. An abundant class of tiny RNAs with probable regulatory roles in Caenorhabditis elegans. Science. 2001;294(5543):858-62. `
12. Lee RC, Ambros V. An extensive class of small RNAs in Caenorhabditis elegans. Science. 2001;294(5543):862-4.
13. Grosshans H, Filipowicz W. Molecular biology: the expanding world of small RNAs. Nature. 2008;451(7177):414-6.
14. Pfeffer S, Zavolan M, Grasser FA, Chien M, Russo JJ, Ju J, et al. Identification of virus-encoded microRNAs. Science. 2004;304(5671):734-6.
15. Gromak N. Intronic microRNAs: a crossroad in gene regulation. Biochem Soc Trans. 2012;40(4):759-61.
16. Wang J, Xiang G, Zhang K, Zhou Y. Expression signatures of intragenic miRNAs and their corresponding host genes in myeloid leukemia cells. Biotechnol Lett. 2012;34(11):2007-15.
17. Sun J, Gao B, Zhou M, Wang ZZ, Zhang F, Deng JE, et al. Comparative genomic analysis reveals evolutionary characteristics and patterns of microRNA clusters in vertebrates. Gene. 2013;512(2):383-91.
18. Zhang Y, Zhang R, Su B. Diversity and evolution of MicroRNA gene clusters. Sci China C Life Sci. 2009;52(3):261-6.
19. Chhabra R, Dubey R, Saini N. Cooperative and individualistic functions of the microRNAs in the miR-23a~27a~24-2 cluster and its implication in human diseases. Mol Cancer. 2010;9:232.
20. Yuan X, Liu C, Yang P, He S, Liao Q, Kang S, et al. Clustered microRNAs' coordination in regulating protein-protein interaction network. BMC Syst Biol. 2009;3:65.
21. Yu J, Wang F, Yang GH, Wang FL, Ma YN, Du ZW, et al. Human microRNA clusters: genomic organization and expression profile in leukemia cell lines. Biochem Biophys Res Commun. 2006;349(1):59-68.
22. Bonci D, Coppola V, Musumeci M, Addario A, Giuffrida R, Memeo L, et al. The miR-15a-miR-16-1 cluster controls prostate cancer by targeting multiple oncogenic activities. Nature medicine. 2008;14(11):1271-7.
23. Griffiths-Jones S. The microRNA Registry. Nucleic Acids Res. 2004;32(Database issue):D109-11.
24. Ambros V, Bartel B, Bartel DP, Burge CB, Carrington JC, Chen X, et al. A uniform system for microRNA annotation. RNA. 2003;9(3):277-9.
25. Lee Y, Kim M, Han J, Yeom KH, Lee S, Baek SH, et al. MicroRNA genes are transcribed by RNA polymerase II. EMBO J. 2004;23(20):4051-60.
26. Cai X, Hagedorn CH, Cullen BR. Human microRNAs are processed from capped, polyadenylated transcripts that can also function as mRNAs. RNA. 2004;10(12):1957-66.
27. Han J, Lee Y, Yeom KH, Nam JW, Heo I, Rhee JK, et al. Molecular basis for the recognition of primary microRNAs by the Drosha-DGCR8 complex. Cell. 2006;125(5):887-901.
28. Yi R, Qin Y, Macara IG, Cullen BR. Exportin-5 mediates the nuclear export of pre-microRNAs and short hairpin RNAs. Genes Dev. 2003;17(24):3011-6.
29. Grishok A, Pasquinelli AE, Conte D, Li N, Parrish S, Ha I, et al. Genes and mechanisms related to RNA interference regulate expression of the small temporal RNAs that control C. elegans developmental timing. Cell. 2001;106(1):23-34.
30. Bartel DP. MicroRNAs: target recognition and regulatory functions. Cell. 2009;136(2):215-33.
31. Gregory RI, Chendrimada TP, Cooch N, Shiekhattar R. Human RISC couples microRNA biogenesis and posttranscriptional gene silencing. Cell. 2005;123(4):631-40.
32. Schwarz DS, Hutvagner G, Du T, Xu Z, Aronin N, Zamore PD. Asymmetry in the assembly of the RNAi enzyme complex. Cell. 2003;115(2):199-208.
33. Brennecke J, Stark A, Russell RB, Cohen SM. Principles of microRNA-target recognition. PLoS Biol. 2005;3(3):e85.
34. Treiber T, Treiber N, Meister G. Regulation of microRNA biogenesis and function. Thromb Haemost. 2012;107(4):605-10.
35. Volinia S, Calin GA, Liu CG, Ambs S, Cimmino A, Petrocca F, et al. A microRNA expression signature of human solid tumors defines cancer gene targets. Proc Natl Acad Sci USA. 2006;103(7):2257-61.
36. Wang Q, Wang S, Wang H, Li P, Ma Z. MicroRNAs: novel biomarkers for lung cancer diagnosis, prediction and treatment. Exp Biol Med (Maywood). 2012;237(3):227-35.
37. Izzotti A, Calin GA, Arrigo P, Steele VE, Croce CM, De Flora S. Downregulation of microRNA expression in the lungs of rats exposed to cigarette smoke. FASEB J. 2009;23(3):806-12.
38. Boeri M, Verri C, Conte D, Roz L, Modena P, Facchinetti F, et al. MicroRNA signatures in tissues and plasma predict development and prognosis of computed tomography detected lung cancer. Proc Natl Acad Sci USA. 2011;108(9):3713-8.
39. Lebanony D, Benjamin H, Gilad S, Ezagouri M, Dov A, Ashkenazi K, et al. Diagnostic assay based on hsa-miR-205 expression distinguishes squamous from nonsquamous non-small-cell lung carcinoma. J Clin Oncol. 2009;27(12):2030-7.
40. Landi MT, Zhao Y, Rotunno M, Koshiol J, Liu H, Bergen AW, et al. MicroRNA expression differentiates histology and predicts survival of lung cancer. Clin Cancer Res. 2010;16(2):430-41.
41. Barshack I, Lithwick-Yanai G, Afek A, Rosenblatt K, Tabibian-Keissar H, Zepeniuk M, et al. MicroRNA expression differentiates between primary lung tumors and metastases to the lung. Pathol Res Pract. 2010;206(8):578-84.
42. Jemal A, Siegel R, Xu J, Ward E. Cancer statistics, 2010. CA Cancer J Clin. 2010;60(5):277-300.
43. Dumont N, Tlsty TD. Reflections on miR-ing effects in metastasis. Cancer Cell. 2009;16(1):3-4.
44. Ma L, Teruya-Feldstein J, Weinberg RA. Tumour invasion and metastasis initiated by microRNA-10b in breast cancer. Nature. 2007;449(7163):682-8.
45. Tavazoie SF, Alarcon C, Oskarsson T, Padua D, Wang Q, Bos PD, et al. Endogenous human microRNAs that suppress breast cancer metastasis. Nature. 2008;451(7175):147-52.
46. Foekens JA, Sieuwerts AM, Smid M, Look MP, de Weerd V, Boersma AW, et al. Four miRNAs associated with aggressiveness of lymph node-negative, estrogen receptor-positive human breast cancer. Proc Natl Acad Sci USA. 2008;105(35):13021-6.
47. Kim K, Chadalapaka G, Lee SO, Yamada D, Sastre-Garau X, Defossez PA, et al. Identification of oncogenic microRNA-17-92/ZBTB4/specificity protein axis in breast cancer. Oncogene. 2012;31(8):1034-44.
48. Tang W, Zhu J, Su S, Wu W, Liu Q, Su F, et al. MiR-27 as a prognostic marker for breast cancer progression and patient survival. PLoS One. 2012;7(12):e51702.
49. Volinia S, Croce CM. Prognostic microRNA/mRNA signature from the integrated analysis of patients with invasive breast cancer. Proc Natl Acad Sci USA. 2013;110(18):7413-7.
50. Karapetis CS, Khambata-Ford S, Jonker DJ, O'Callaghan CJ, Tu D, Tebbutt NC, et al. K-ras mutations and benefit from cetuximab in advanced colorectal cancer. N Engl J Med. 2008;359(17):1757-65.
51. Van Cutsem E, Kohne CH, Hitre E, Zaluski J, Chang Chien CR, Makhson A, et al. Cetuximab and chemotherapy as initial treatment for metastatic colorectal cancer. N Engl J Med. 2009;360(14):1408-17.
52. Lievre A, Bachet JB, Boige V, Cayre A, Le Corre D, Buc E, et al. KRAS mutations as an independent prognostic factor in patients with advanced colorectal cancer treated with cetuximab. J Clin Oncol. 2008;26(3):374-9.
53. Johnson SM, Grosshans H, Shingara J, Byrom M, Jarvis R, Cheng A, et al. RAS is regulated by the let-7 microRNA family. Cell. 2005;120(5):635-47.
54. Chen X, Guo X, Zhang H, Xiang Y, Chen J, Yin Y, et al. Role of miR-143 targeting KRAS in colorectal tumorigenesis. Oncogene. 2009;28(10):1385-92.
55. Tsang WP, Kwok TT. The miR-18a* microRNA functions as a potential tumor suppressor by targeting on K-Ras. Carcinogenesis. 2009;30(6):953-9.
56. Mekenkamp LJ, Tol J, Dijkstra JR, de Krijger I, Vink-Borger ME, van Vliet S, et al. Beyond KRAS mutation status: influence of KRAS copy number status and microRNAs on clinical outcome to cetuximab in metastatic colorectal cancer patients. BMC Cancer. 2012;12:292.
57. Mosakhani N, Lahti L, Borze I, Karjalainen-Lindsberg ML, Sundstrom J, Ristamaki R, et al. MicroRNA profiling predicts survival in anti-EGFR treated chemorefractory metastatic colorectal cancer patients with wild-type KRAS and BRAF. Cancer Genet. 2012;205(11):545-51.
58. Svoboda M, Sana J, Fabian P, Kocakova I, Gombosova J, Nekvindova J, et al. MicroRNA expression profile associated with response to neoadjuvant chemoradiotherapy in locally advanced rectal cancer patients. Radiat Oncol. 2012;7:195.
59. Krutzfeldt J, Rajewsky N, Braich R, Rajeev KG, Tuschl T, Manoharan M, et al. Silencing of microRNAs in vivo with 'antagomirs'. Nature. 2005;438(7068):685-9.
60. Garzon R, Marcucci G, Croce CM. Targeting microRNAs in cancer: rationale, strategies and challenges. Nat Rev Drug Discov. 2010;9(10):775-89.
61. Ebert MS, Neilson JR, Sharp PA. MicroRNA sponges: competitive inhibitors of small RNAs in mammalian cells. Nat Methods. 2007;4(9):721-6.
62. Esau CC. Inhibition of microRNA with antisense oligonucleotides. Methods. 2008;44(1):55-60.
63. Ebert MS, Sharp PA. MicroRNA sponges: progress and possibilities. RNA. 2010;16(11):2043-50.
64. Pereira DM, Rodrigues PM, Borralho PM, Rodrigues CM. Delivering the promise of miRNA cancer therapeutics. Drug Discov Today. 2013;18(5-6):282-9.
65. Branch AD, Rice CM. Antisense gets a grip on miR-122 in chimpanzees. Science Translational Medicine. 2010;2(13).
66. Jopling CL, Yi M, Lancaster AM, Lemon SM, Sarnow P. Modulation of hepatitis C virus RNA abundance by a liver-specific MicroRNA. Science. 2005;309(5740):1577-81.
67. Vester B, Wengel J. LNA (locked nucleic acid): high-affinity targeting of complementary RNA and DNA. Biochemistry. 2004;43(42):13233-41.
68. Weber JA, Baxter DH, Zhang S, Huang DY, Huang KH, Lee MJ, et al. The microRNA spectrum in 12 body fluids. Clin Chem. 2010;56(11):1733-41.
69. Turchinovich A, Weiz L, Langheinz A, Burwinkel B. Characterization of extracellular circulating microRNA. Nucleic Acids Res. 2011;39(16):7223-33.
70. Allegra A, Alonci A, Campo S, Penna G, Petrungaro A, Gerace D, et al. Circulating microRNAs: new biomarkers in diagnosis, prognosis and treatment of cancer (review). Int J Oncol. 2012;41(6):1897-912.
71. Gu L, Song G, Chen L, Nie Z, He B, Pan Y, et al. Inhibition of miR-21 induces biological and behavioral alterations in diffuse large B-cell lymphoma. Acta Haematol. 2013;130(2):87-94.
72. miRInform Pancreas [Internet]. Austin, TX: Asuragen Inc. c2013 [2013 Oct 17]. Available from: http://asuragen.com/products-and-services/clinical-lab/mirinform-pancreas/.
73. Szafranska AE, Davison TS, John J, Cannon T, Sipos B, Maghnouj A, et al. MicroRNA expression alterations are linked to tumorigenesis and non-neoplastic processes in pancreatic ductal adenocarcinoma. Oncogene. 2007;26(30):4442-52.
74. Szafranska AE, Doleshal M, Edmunds HS, Gordon S, Luttges J, Munding JB, et al. Analysis of microRNAs in pancreatic fine-needle aspirates can classify benign and malignant tissues. Clin Chem. 2008;54(10):1716-24.
Received in October, 2013.
Accepted in March, 2014.
Alejandro Moro-Soria. Instituto Nacional de Oncología y Radiobiología, INOR. Calle 29 y F, CP 10400, Plaza de la Revolución, La Habana, Cuba. E-mail: alexmoro@infomed.sld.cu

{kind=link}