










My SciELO
 Custom services
Custom servicesServices on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkBiotecnología Aplicada
On-line version ISSN 1027-2852
Biotecnol Apl vol.32 no.1 La Habana Jan.-Mar. 2015
REVIEW
Pharmacological applications and in vitro biotechnological production of anticancer alkaloids of Catharanthus roseus
Aplicaciones farmacológicas y producción biotecnológica in vitro de los alcaloides anticancerígenos de Catharanthus roseus
Hebert Jair Barrales-Cureño
Instituto de Genética. Colegio de Postgraduados Campus Montecillo. Km. 36.5 Carret. Mexico-Texcoco. Montecillo, Texcoco, State of Mexico 94420, Mexico.
ABSTRACT
Catharanthus roseus (periwinkle) is a plant of the Apocynaceae family. This plant synthesizes two indole terpene alkaloids: vinblastine and vincristine, used against cancer. C. roseus is endemic to Madagascar and is known as vicaria in Cuba; in Mexico, it is found in Veracruz, Tabasco, Quintana Roo and Yucatan. C. roseus is of considerable interest for in vivo or in vitro studies, and from which over 130 alkaloids of the indole terpene group have been isolated. The total alkaloid content in roots is 2-3 %, 9 % in the fibers, the leaves contain 1 % alkaloids, the stem, fruit, seeds, and pericarp containing 0.48, 0.40, 0.18 and 1.14 %, respectively. About 500 kg of dried leaves are used to isolate 1 g of vinblastine and two tons of macerated leaves provide 1 g of the active principle required for the treatment of a child with leukemia for 6 weeks. Semi-synthesis and organic synthesis are costly and low yielding, so alternatives have been used to enhance biotechnological production by adding inducers to stimulate the production of metabolites in the biosynthetic pathway. Therefore, the objectives of this paper are: to mention the biological mechanism of action, biological activity, biosynthetic pathway of the vinblastine and vincristine alkaloids, report on the production of alkaloids from in vitro callus cultures and cells in suspension. Finally, the methods for the quantification of terpene indole alkaloids are highlighted.
Keywords: terpene indole alkaloids, mechanism of action, anti-leukemia drugs, vinblastine, vincristine, in vitro culture.
RESUMEN
Aplicaciones farmacológicas y producción biotecnológica in vitro de los alcaloides anticancerígenos de Catharanthus roseus. Catharanthus roseus es una planta que pertenece a la familia Apocynaceae. Esta planta sintetiza dos alcaloides indol-terpénicos cuyo uso es relevante para el tratamiento contra el cáncer: vinblastina y vincristina. C. roseus es endémica de Madagascar y se conoce como vicaria en Cuba; en México, se distribuye en Veracruz, Tabasco, Quintana Roo y Yucatán. Es de gran interés para estudios in vivo o in vitro, y de ella se han aislado más de 130 alcaloides del grupo indol-terpénico. Su contenido total de alcaloides en raíces es de 2-3 % (9 % en sus fibras), las hojas contienen 1 % de alcaloides, el tallo, fruto, semillas, pericarpio contiene 0.48; 0.40; 0.18 y 1.14 %, respectivamente. Se utilizan cerca de 500 kg de hojas secas para aislar 1 g de vinblastina y dos toneladas de hojas maceradas proporcionan 1 g del principio activo, cantidad requerida para el tratamiento de un niño con leucemia durante 6 semanas. La semisíntesis y la síntesis orgánica son costosas y de baja producción, por lo que se han utilizado alternativas biotecnológicas para incrementar la producción mediante la adición de inductores que estimulan la producción de los metabolitos en su ruta de biosíntesis. En este trabajo se abordan el mecanismo de acción y las actividades biológicas de esta planta, la ruta de biosíntesis de los alcaloides vinblastina y vincristina, y la producción de alcaloides a partir de cultivos in vitro de callos y células en suspensión. Finalmente, se destacan los métodos de cuantificación de los alcaloides indol-terpénicos.
Palabras clave: alcaloides indol-terpénicos, mecanismo de acción, fármacos antileucémicos, vinblastina, vincristina, cultivo in vitro.
INTRODUCTION
Catharanthus roseus L. (G.) Don., is an important medicinal plant belonging to the Apocynaceae family; this plant is a dicotyledonous angiosperm and synthesizes two terpene indole alkaloids: vinblastine and vincristine that are used to fight cancer [1]. The plant species C. roseus is endemic to Madagascar and is commonly known in Cuba as vicaria; it is found in Mexico in the States of Veracruz, Tabasco, Quintana Roo and Yucatán [2] as well as in El Salvador [3]. This plant is also found in other tropical and subtropical regions such as: South Africa, Southern Asia, South America and Australia where it is used as an ornamental plant; it grows well in India, Israel, Sri-Lanka, Mozambique [4] and Egypt [1]. In Cuba, it is frequently grown by the population in yards and gardens as an ornamental and medicinal plant [5].
The C. roseus species is of great interest for in vivo or in vitro studies from which more than 130 alkaloids of the terpene indole group and 25 alkaloids of dimeric nature have been isolated [6-8]; several of them are used in human medicine. All parts of the plant contain more than 100 alkaloids in variable proportions. The total alkaloid content of roots reach 2-3 % or nearly 9 % in root fibers, while the leaves contain 1 % alkaloids, the stem, fruit, seeds, and pericarp contain 0.48, 0.40, 0.18 and 1.14 %, respectively [9]. This plant produces anti-cancer dimeric alkaloids
such as vinblastine and vincristine in concentrations of 0.0004 to 0.0003 % of the dry weight of leaves and stores anti-hypertensive alkaloids such as ajmalicine and serpentine in the roots. Certain chemical constituents are vinblastine, vindoline, catharantine, ajmalicine and serpentine [8]; the species are also rich in bisindole alkaloids (approximately 40 compounds), many of which contain a molecule of vindoline or catharantine [8]. The market for its leaves is monopolized by the United States, and by a country in Eastern Europe: Hungary. The main demand for leaves is by the United States, and the United Kingdom is also interested in buying 10 tons of leaves each year and Germany shows great interest in the roots of this plant [3].
The biosynthetic route for indole alkaloids has been studied by De-Luca and Cutler [10]. Because of the presence of cytotoxic bis-indole alkaloids of therapeutic importance, the production of vinblastine, vincristine and vindesine has become one of the main fields of interest in modern cell biotechnology [11]. The plant produces the active dimeric alkaloids in low concentrations (0.0005 %), where nearly 500 kg of dry leaves of C. roseus are used to isolate 1 g of vinblastine [8] and 2 tons of macerated leaves produce 1g of the alkaloid as the active principle, which is the amount required for the treatment of a child with leukemia for 6 weeks [9]. Because of the large number of alkaloids it contains, the isolation of vinblastine and vincristine in the laboratory is very costly.
Although all parts of the plant produce alkaloids (leaves and stems) in different proportions [12], the maximum concentrations are found in the cortex of the roots, particularly when blooming [13]. A wide array of different alkaloid sub-classes have been identified: vincosan, corynanthean, vallesiachotaman, strychnan, aspidospermatan, plumeran, ibogan, eburnan and bisindole alkaloids [14]. Up to 40 different bis-indole alkaloids have been found in C. roseus, many of which contain a moiety of plumerane (vindoline) and ibogane (catharantine). In relation to plant chemistry, C. roseus contains carbohydrates, flavonoids, saponins, phenol compounds, terpene indole alkaloids [15], antocyanines, glucosides [16], heart glycosides, steroids, mono-terpene glucosides [8]. It is also has no tannins. Two flavonols have also been isolated and identified [17] as well as glycosidic flavonols that have been identified in seeds, stems, leaves and flowers of C. roseus [18]. The extracts of the sprouts of C. roseus are used as a potential source of natural available antioxidants and with excellent pharmaceutical applications [19].
The biological mechanism of action of the vinblastine and vincristine consists of the binding of the tubulin during mitosis. These compounds inhibit the chromatin filaments drawn to their respective poles [20], leading to the inhibition of cellular mitosis during the metaphase, and thereby starting the programmed cellular death or apoptosis [21]. Vincristine inhibits polymerization of the microtubules, producing an arrest in phase G2/M and inducing apoptosis [22].
Because the semi-synthesis of vincristine and vinblastine, starting with the precursors and the organic synthesis, is highly expensive and the production is poor, and also the extraction of the metabolites are complicated by the low concentrations of the pharmaceuticals in C. roseus, alternative biotechnology strategies have been used to be able to increase the production of these secondary metabolites. They include the addition of biotic or abiotic inducers that stimulate the production of the metabolites in the biosynthesis pathway of the alkaloids. Therefore, the aims of this paper are: to mention the biological mechanism of action, biological activities and the biosynthetic pathways for the vinblastine and vincristine alkaloids, to inform on the production of alkaloids through in vitro cultures of calluses and cells in suspension, and finally on the quantification methods of the terpene indole alkaloids.
ON THE RELEVANCE OF C. roseus ALKALOIDS FOR ANTICANCER THERAPY
Particularly, the use of C. roseus as source of anticancer drugs has become relevant due to the high morbidity and mortality rates of certain types of cancers, which are sensitive to treatment with compounds like vinblastine and vincristine. This is the case of acute myeloid leukemia [23] and Hodgkin’s lymphoma.
To have an idea of the possible impact of these therapies, there have been reported 350 434 cases of leukemia in the world, with a standard incidence rate both in underdeveloped and developed countries of 5.0 per 100 000 inhabitants [24], accounting for approximately 210 000 cases in the last ones. The highest rates have been found in the United States, Canada, Europe and Australia; with the lowest in African countries [25]. Moreover, acute myeloid leukemia has a peak incidence in the group of children of less than five years of age, descending in the group of 5 to 9 years of age and as of that age it increases exponentially with age [26]. This makes the availability of such compounds more urgent for the scientific and medical community.
In relation to Hodgkin’s Lymphoma, it has an incidence of 3 cases per 100 000 yearly, and comprises 10 % of the lymphomas in the United States, of which 85 % are found in males, with a bimodal incidence curve: 15 to 34 years of age and after 50 years of age. Estimates of the American Cancer Association in 2010 established that there were 8490 cases of which 4670 were men, with 1320 deaths [27]. In the European Union, the incidence is of 2.2 per 100 000/year, with mortality rates of 0.7 per 100 000/year [28]. Noteworthy, during the last decade, the survival of patients treated for Hodgkin’s Lymphoma substantially improved, and the percentage of cure for this neoplasia is of 80 to 85 % [29].
Particularly in Mexico, until 2003 there were 935 cases reported, with a higher incidence in the group of males of 15 to 19 years of age and females had the same incidence in the groups of 15 to 19 and 20 to 24 years of age [24]. In fact, according to the INCan, the Mexican National Institute of Cancer, until 2004 it represented 0.8 % of the lymphomas, with 162 cases diagnosed, of which 88 were men and 74 were women [30, 31].
All these have fostered the search for new compounds and the development of efficient production processes for its obtainment, using C. roseus as biological source for alkaloids displaying therapeutic activity relevant for cancer treatment.
MECHANISMS OF BIOLOGICAL ACTION OF THE ALKALOIDS OF C. roseus
The cytotoxic alkaloids of C. roseus (vinblastine, vincristine and more recently, vinorelbine) are anti-mitotic, anti-cancer agents inducing tubuline to form spiral polymers at physiological protein concentrations [32] thus interfering in the formation of microtubules.
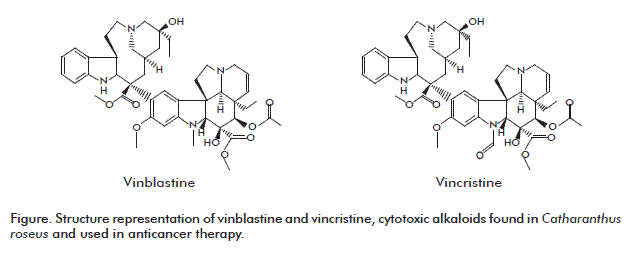
Vinblastine is an analogous chemical compound of vincristine (Figure). It binds to the tubulin and thus interferes in the assembly of the microtubules [33].
During mitosis metaphase, the pharmaceutical inhibits polymerization of the tubulin to microtubules, in contrast to the anti-cancer drug, taxol, which has the function of stabilizing the microtubules, reducing their dynamism by later impeding depolymerization and promoting the arrest of the mitosis, and therefore producing, cell death [34].
The vinblastine works specifically during the metaphase stage of the cell cycle, since the microtubules are components of the mitotic spindle, and the kinetochores that are necessary for the separation of the chromosomes during the anaphase in mitosis. Vincristine enters the cell through a transportation mechanism and it binds to the tubular proteins.
The prescribed dosages of vinblastine range from 3.7 to 18.5 mg/kg with an interval of at least seven days between each dose; having a half clearance life of 25 hours and 95 % of it is excreted in the feces and less than 1 % is eliminated in the urine without metabolization [23].
Vincristine is mainly metabolized in the liver. Its main metabolite is the diacetylvinblastine, with a greater biological activity [35]. It also interferes with the nucleic acid and protein synthesis by blocking the use of glutamic acid. Other effects in the DNA and RNA syntheses were recently described, also in the inhibition of the proteosome [36], anti-angiogenesis [37] and the decrease of the resistance of the cells to chemotherapy [38].
Furthermore, the vincristine sulfate is used to treat Wilm’s tumor, neuroblastoma, breast cancer, rabdomyosarcoma and osteogenic sarcoma.
USES OF C. roseus IN TRADITIONAL MEDICINE
Traditional medicine uses the alkaloids from C. roseus for different non-malignant diseases: in Africa the leaves are used for menorrhagia and rheumatism [39].
C. roseus has been used to fight diabetes for many years; it was proven by the induction of diabetes in rats that it had hypoglycemic activity [40], on having an anti-diabetic effect as the result of the increase of glucose and the promotion of insulin production [41, 42].
In India, wasp bites are treated with the sap from the leaves, and in Brazil it is used against diabetes, in the treatment of hemorrhages and wound healing. In Hawaii an extract of the boiled plant has been prescribed to stop bleeding. In Central and South America, they use it to relieve throat pain and laryngitis. In Cuba, Puerto Rico, Jamaica and other islands, the aqueous extract of the white flower is commonly used as an eye wash for infants and as fomentation for easing eye diseases [39, 43].
In the Bahamas they use the infusion of flowers for asthma, and of the entire plant to fight tuberculosis. In Mauritius, the infusion of leaves is used to combat dyspepsia and indigestion. In Vietnam, it is used for malaria and diabetes; and the inhabitants of Bermuda and Curacao use the plant when they have high blood pressure [44].
It has been also used for treating fever, malaria, menstrual cycle regulation, as a euphoric drug [45], as tranquilizer, and for its ability to reduce arterial blood pressure [42].
The plant is also known for its anti-spasmodic properties because of the presence of alkaloids such as ajmalicine, serpentine and reserpine. The alkaloids: vincamine and vindoline are anti-ulcer compounds. In France, the hot water extract of the whole plant is used as an anti-galactogogue [46].
Finally, C. roseus shows anti-spasmodic properties because of its reserpine and serpentine contents. Furthermore, it shows high anti-plasmodial activity in vitro because of the presence of terpenoids, flavonoids and sesquiterpenes [47]. C. roseus should not be consumed orally without cooking because it may be hallucinogenic [47, 48].
USES OF C. roseus IN ALLOPATHIC MEDICINE
The anti-tumor alkaloids vinblastine and vincristine are used in malignant diseases; they are used in chemotherapy for leukemia since they reduce the number of leukocytes in the blood (a high number of leukocytes indicate leukemia) and in the treatment of Hodgkin’s disease [13] characterized by being a monoclonal B cell neoplasia, and by the presence of abnormal cells called Reed-Sternberg cells [49].
Vinblastine (vinblastine sulfate) is experimentally used for neoplasia treatment and for resistant pregnancy choriocarcinoma, a malignant neoplasia of the trophoblast, which is a highly aggressive and fetal lesion, since even when there is a timely diagnosis and it is appropriately treated with chemotherapy, it produces mortality in 10 to 15 % of the cases [50].
It is also effective in the treatment of advanced testicular tumors, breast cancer, Kaposi sarcoma, and the Letterer-Siwe disease [51]. Vincristine, formally known as leurocristine (vincristine sulfate) is used in leukemia treatment in children.
Vincristine is produced by the bonds of the terpene indole alkaloids: vindoline and catharantine in the C. roseus plant [52]. The use of the vinblastine and vincristine combined with chemotherapy has given 80 % remission in Hodgkin’s disease, 99 % in acute lymphocytic leukemia, 80 % in Wilm’s tumor in children, 70 % in pregnancy corium cancer and the remission of 50 % in Burkitt’s lymphoma [39, 43]. The indole alkaloid called amotin also has a strong anti-leukemia activity [1].
At the neurologic level, the ajmalicine and serpentine are drugs used in treating depression, anxiety, as well as being effective as anti-stress drugs [53, 54]. The supplements based on active ingredients of C. roseus such as vincamine, are used for the prevention and treatment of cerebro-vascular disorders and failures, vertigo, ischemic deficiencies and headaches, because they help oxygenate and increase brain glucose levels [55]; besides preventing abnormal
clotting, they also increase the levels of serotonin, a brain neurotransmitter. It is important to mention that the deficiency of serotonin produce schizophrenia, phobia, migraine and bulimia.
On the other hand, vincamine is now known to increase the memory retention properties and it is effective in the treatment of vascular dementia [56]. Anhydrous vinblastine is used in the treatment of lung and cervix cancer [57]. The catharantine isolated from C. roseus is cytotoxic in P-388 and KB human cancer cell lines [58]. Furthermore, ajmalicine is used in the treatment of circulatory disorders and as an anti-hypertensive since it acts as an antagonist of the α1-adrenergic receptor, known as alpha blocker [59], with a preferential action on α2-adrenergic receptors [60].
BIOLOGICAL ACTIVITIES OF THE C. roseus ALKALOIDS
The main secondary metabolites of C. roseus are terpene indole alkaloids with important applications in human medicine as mentioned above, and also presenting biological activities such as: anti-tumor, anti-diabetes, anti-helminthic, anti-hypertensive, anti-diarrhea, and anti-microbial actions, among others. The clinical evidence of the biological activities of C. roseus is presented below.
Anti-tumoral activity
Cancer is a genetic pathology characterized by the uncontrolled proliferation of a certain group of cells in the body. If the control regulating cell multiplication does not work well, the cells start growing and dividing unnecessarily, and when the young cells inherit the trend to proliferate without any control, the result is a clone that expands indefinitely forming a tumor.
These tumors may be benign, or they may become malignant if they are able to invade and disseminate throughout the body, in a process known as metastasis [61].
Different percentages of crude methanol extracts of Catharanthus have shown anti-cancer activity against many types of cells under in vitro conditions [62] and a greater activity is especially found against multi-drug resistant tumors [63].
As early as in 1979, ethanol extract from leaves (70 %) was found as very active in CA-Ehrlich ascitis when administered intra-peritoneally in female mice [64]. The alkaloid fraction of the dry leaves is also active at surface level. Nineteen patients with genital, flat and common warts were treated; of these, all warts disappeared in six patients, almost completely in seven patients, 50 % of the warts were eliminated in five patients and one patient did not show any response to the chloroform extract [65].
The chloroform extract of leaves was active in Leuk-P3887 [66]. The total amount of alkaloids of the whole plant were administered to mice by the intra-peritoneal route at a dose of 10 mg/kg and by the oral route at 75 mg/kg, both concentrations found to be active in Leuk-P15348.
Anti-helminthic activity
The infections produced by helminths are chronic diseases affecting human beings. C. roseus is used as an anti-helminthic agent in medicine.
Agarwal et al. evaluated the anti-helminthic property of C. roseus through the use of Pherethima posthuma as the experimental model with piperazine citrate as the standard reference. The ethanol extract, at a concentration of 250 mg/mL showed significant anti-helminthic activity at 46.3 min, while the standard drug showed activity at a concentration of 50 mg/mL at 40.7 min. This ethno-medical research considers C. roseus as an effective anti-helminthic drug [67].
Anti-diarrheal activity in vivo
The in vivo anti-diarrheal activity of the ethanol extract of leaves has been tested in Wistar rats, using castor oil as the experimental diarrhea inducing agent, besides the pre-treatment of the extract. Loperamide and the atropine sulfate were used as the standard drug. The anti-diarrhea effect of the ethanol extract of C. roseus showed the dose dependent inhibition of the diarrhea inducing castor oil at a dose of between 200 and 500 mg/kg [68].
Anti-oxidant activity
Kumar et al., 2012 studied the anti-oxidant activity of C. roseus. These researchers found that the temperature of the habitat had a specific effect on the antioxidant activity, which would make it possible to see that the super oxide dismutase and polyphenol oxidase enzymes had a greater antioxidant activity when the temperature was increased in contrast to the catalase [69].
The methanol extract of the leaf was used to direct the study. Rasool et al., in 2011 carried out several in vitro tests of antioxidants to study the effect of the solvent on the extraction and total anti-radical potential of the different extracts of C. roseus. The trials showed that the extracts and the fractions are a good source of natural antioxidants. The 100 % methanol extract and the fraction of 100 % ethyl acetate of the shoots of C. roseus showed high antioxidant activity [70].
Antimicrobial activity
Muhammad et al. reported the anti-bacterial potential in crude extracts of different parts (leaves, stalks, roots and flowers) of C. roseus against clinically relevant bacterial strains [71]. The antimicrobial activity of extracts of leaves of C. roseus was tested against microorganisms such as Pseudomonas aeruginosa NCIM 2036, Salmonella typhimurium NCIM 2501 and Staphylococcus aureus NCIM 5021 and it was found that the crude extract may be used as a prophylactic agent in the treatment of many diseases [72].
Kumari and Gupta evaluated the potential of C. roseus against several pathogenic germs under in vitro conditions, at the dose of 50 mg/mL of the extract of C. roseus var. rosea which was effective against Bacillus fusiformis, while at the dose of 20 mg/mL of the extract it was selective against Aspergillus fumigatus, Candida albicans, Escherichia coli, Bacillus fusiformis in two varieties of C. roseus: Rosea and Alba [42].
The ethanol extract of flowers of C. roseus is reported to have healing properties in rats [73]. The extract promotes contraction of wounds, increases the content of hydroxyproline and the anti-bacterial activity against P. aeruginosa and S. aureus.
Furthermore, the ethanol extract of leaves, stems, flowers and root extracts of C. roseus have a wide scope of anti-bacterial activity against E. coli, Streptococcus pyogenes, Streptococcus agalactiae, Salmonella typhi and Aeromonas hydrophila [71]. In a similar study, the extracts of different parts of the plant were reported to inhibit both the gram-positive and gram-negative bacteria [74].
Cytotoxic activity
The alkaloid fraction of dry leaves in a cell culture was active in CA-9KB, the median effective dose (ED50) at 0.0435 µg/mL [75]. The chloroform extract and filtrate of the in vitro culture of calluses, at the dose of 50 mg (dry weight of the plant) was active on the aqueous extract of the culture Leuk-L12. The methanol extracts of the leaves of C. roseus produce positive anti-proliferative activity against HeLa human cancer cells (a HeLa cell is a cell type classified in an immortal cellular line used in scientific research. Its growth is aggressive, and its resistance to apoptosis is mainly due to a combination of papilloma virus 18 that produces a protein that degrades p53 without mutating it, and several alterations in chromosomes 1, 3, 5 and 6), MCF-7 (it is an epithelial cancer cell line that has been widely used, and it is derived from breast adenocarcinoma) with values of the concentration showing 50 % of maximum inhibition of cell proliferation (GI50) of 3.5 ± 0.1 and 4.7 ± 0.6 μg/mL, respectively [76].
Anti-hyperglycemic effect
The daily oral administration of the extract of dichloromethane:methanol (1:1) has been evaluated by using the leaves of C. roseus (500 mg/kg of body weight) for 20 days and its effect was tested in blood glucose and normal liver enzymes in diabetic rats [77]. The extract showed a significant increase in the body weight and a decrease in glucose, urea and cholesterol levels of the treated animals. The activity of the liver enzymes such as hexokinase, was increased while that of glucose-6 phosphatase and fructose 1,6-biphosphatase decreased significantly.
Anti-diabetic effect
Several studies in animals have shown that the ethanol extracts of leaves and flowers of C. roseus decrease the levels of glucose in the blood [78, 79]. Furthermore, the aqueous extracts have the capacity to decrease the blood glucose in 20 % in trials with diabetic rats, while the reductions of the levels of glucose in the blood with dichloromethane and methanol extracts are of 49 and 58 %, respectively [80].
On the other hand, the vindolicine alkaloid demonstrated a potent inhibition activity in PTP-1B, which is due to the effect of vindolicine as a new inhibitor of PTP-1B, which can serve as a “sensitizer of insulin” in the management of type 2 diabetes. Para-nitrophenyl phosphate (pNPP) was used as a substrate for the trial on phosphatase activity (the recombinant enzyme PTP-1B) and it was added at the start of the reaction [81].
The main structural skeleton of the terpene indole alkaloids together with its biological activity are shown in table 1.
GENERAL BIOSYNTHESIS OF THE ALKALOIDS OF C. roseus
The secondary metabolic route of the indole alkaloids in C. roseus has been documented [82, 83] and it is very complex. The biosynthesis route of the indole alkaloids has been studied by De-Luca and Cutler [10]. The tryptophan (precursor of the indoles) turns into a tryptamine, and later it is condensed with secologanin (a precursor of the iridoids) to form strictosidine, the common precursor of all indole alkaloids, divided into three branches of several enzymatic reactions which leads to the production of ajmalicine, tabersonine and catharanthine, respectively. The transformation of vindoline, a downstream enzymatic intermediary product of tabersonine, into the downstream processing pathway of catharantine followed by several enzymatic reactions generates vinblastine, which is finally transformed in a single reaction to vincristine. Notably, the production of terpene indole alkaloids is strongly regulated by environmental conditions and the cell growth stage [84, 85].
PRODUCTION OF ALKALOIDS IN IN VITRO CULTURES OF C. roseus CALLUSES
The in vitro culture technique of tissues and cells is considered to be an effective tool for the production of secondary metabolites. The in vitro culture of plant cells, whether this is done in a solid medium with calluses or in a liquid medium with cell suspensions, is a potential source of substances of interest for the pharmaceutical industry [86].
The in vitro cultures of calluses are obtained by explants of leaves of different plant species [87] and by obtaining in vitro cultures of viable cells in suspension that are obtained through the inclusion of friable calluses (with disintegration capacity) in liquid culture media.
Current research in relation to obtaining in vitro cultures of calluses and cells from C. roseus is presented below. Fatemeh and Abdolreza [88], obtained the maximum induction of calluses as well as a high percentage of fresh and dry weights of calluses obtained from explants of roots with 1.5 m/L of naphthalene acetic acid (NAA) and 0.1 mg/L from kinetin and 2 g/L of glutamic acid under conditions of darkness. Haq et al. [89] implemented an efficient protocol of the micropropagation through explants, from nodal portions, shoots and callus-genesis by means of explants of leaves and knots [89]. There were multiple outgrowths obtained from several concentrations of benzyl amino purine (BAP) and NAA, but BAP (1 mg/L) showed the best responses (90 and 80 %) of both explants. The best response of calluses was observed in the Murashige-Skoog (MS) medium supplemented with 2, 4-D plus Kin (1 + 1 mg/L) in all explants (the result of obtaining 95 % leaves, 80 % nodes) and the plantelets obtained were transferred to different pots under greenhouse conditions for their adaptation.
Soleimani et al. [12] indicated that the callus cultures with applications of 2,4-D have a significant effect on the percentage of callus-genesis and on the amounts of vinblastine and vincristine alkaloids [12], but the kinetin in high concentrations produces a considerable decrease in the percentage of callus and alkaloid production [12]. The maximum concentration of vincristine (0.7088 µg/g of dry weight) was produced with 1.0 mg/L of the 2,4-dichlorophenoxi acetic acid (2,4-D plus 0.5 mg/L of kinetin and the best treatment for the maximum production of vinblastine (0.7088 µg/g of dry weight) was obtained with 1.0 mg/L of 2,4-D without kinetin. Verma et al. [90] obtained an early induction of calluses (99 %) on the tenth day through the use of explants of hypocotyl under conditions of darkness in the MS medium, where the best one was that containing: benzyl aminopurine (BAP 1.5 mg/L) plus NAA (1.0 mg/L), followed by the MS medium fortified with BAP (3.0 mg/L) plus NAA (4.0 mg/L). The MS medium with half the content of macro- and micro-nutrients supplemented with indole butyric acid (IBA; 2.5 mg/L) plus NAA (0.5 mg/L) showed the best response in rooting with high quality roots [90].
Verma et al. [91] analyzed combinations of auxins and cytokinins to find the best callus growth and increase the alkaloid content. The highest content of total alkaloids found in the production of the biomass of calluses was through the leaves having the concentrations of 0.50 mg/L 2,4-D and 1.0 mg/L BAP and 6 % sucrose. Negi [92] suggested a simple protocol to obtain a good production of calluses from the nodal explants of C. roseus: an MS culture medium supplemented with a combination of 1.0 mg/L kinetin and a combination of 1.0 mg/L BAP; 1 mg/L 2,4-D and 1 mg/L IAA [92]. Similarly, MS culture media supplemented with a combination of 2 mg/L kinetin and BAP; MS plus 2, 4-D, IAA and 0.5 mg/L showed green calluses and resin secretion. However, the MS culture media plus 2 mg/L of BAP and 1 mg/L of 2, 4-D produced better calluses of a green color with resin secretion. Furthermore, the leaves explants growing in the MS medium plus 1.0 mg/L of BAP and 1.0 mg/L of NAA showed a large number of roots. When combining in the MS medium, 0.1 mg/L of kinetin and 1.0 mg/L of 2,4-D; and MS plus 0.1 mg/L BAP and 1 mg/L IAA, a simple process for the induction of C. roseus calluses was observed.
On the other hand, Kalidass et al. [93], established in vitro cultures of calluses of C. roseus in MS media supplemented with 1.0 µM of 2,4-D and 1.0 µM of 6-furfurylaminopurin to grow callus cultures, and they obtained 598.04 mg of dry biomass to quantify the content of vincristine by HPLC through the use of methanol extracts. The concentration of 0.5 µM of benzyl adenine (BA) produced 0.57 mg/g dry weight of vincristine while the concentration of 1.0 µM NAA and 0.5 µM BA produced 20.38 mg/g of dry weight of vincristine. Ataei-Azimi et al. studied the production of indole alkaloids in in vitro cultures of calluses and roots. The alkaloids came from calluses and roots of petioles of C. roseus in the presence of 0.1, 5, 10 and 20 mg/L of kinetin and NAA [15]. The MS medium with 0.1 mg/L NAA plus 0.1 mg/L of kinetin showed the highest production of vindoline, catharantine, vincristine and the capacity of the organogenesis of roots, but the levels of those alkaloids and of ajmalicine were low in comparison with the petioles of the whole plant and the levels of serpentine were similar. Those results suggest that the capacity to synthesize vinblastine and vincristine, vindoline and catharantine are associated to morphologic differentiation, light and high temperature (35 ºC).
New roots, callus roots and calluses from the MS medium were obtained with 0.1 mg/L of NAA plus 0.1 mg/L of kinetin and were sub-cultured in a hormone free medium with the same hormonal concentrations for their growth and organogenesis. The best treatment for the formation of calluses was with 2 mg/L BAP plus 2 mg/L kinetin and 0.1 mg/L indole acetic acid (IAA) [12]. Ferderique and Leslie [94] reported that cytokinins have a greater accumulation effect than the indole alkaloids such as ajmalicine and serpentine in cultures of calluses of C. roseus taken from cotyledons.
Previous studies show that adding the precursors of the biosynthesis of alkaloids may be a production strategy for the secondary metabolites. The treatment of tryptophan on calluses of C. roseus can increase the content of catharantine up to 950.536 μg/g of dry weight [95]. Generally, the plants with poor callus growth produce more catharantine than those of greater growth. From such a study it was found that the optimum growth was observed with the treatment of 175 mg/L of the tryptophan precursor.
Finally, some alkaloids that have been recently discovered and isolated from in vitro cultures of calluses of C. roseus: akuammicine; cavincine; tubotaiwine; catharantine; tabersonine; perivine; 21-hydroxy cyclolochnerine; lochneridine; alstonine; serpentine; vallesiachotamine; isovallesiachotamine; ajmalicine; 19-epi,3-iso ajmalicine; 3-epi ajmalicine; O-diacetyl akuammiline; tetrahidroalstonine; sitsirikine; yohimbine; dihydro-sitsirikine,; 7-hydroxiindolenine ajmalicine; pseudo-indoxil ajmalicine; 10-hydroxydeacetyl akuammiline; mitraphilline; strichtosidine; vinblastine and two previously unidentified compounds: cavincidine and perosine [8].
PRODUCTION OF ALKALOIDS IN in vitro CELL CULTURES OF C. roseus
The in vitro suspension cell cultures of C. roseus are used to study the structural modifications of several synthetic and natural products [96] such as oxidation, hydroxylation, reduction, isomerization, esterification and glycosylation [97], as well as obtaining the maximum production of secondary metabolites of medical importance. C. roseus calluses and cells have been studied for their biotransformation capacity [98]. The increase of the accumulation of ajmalicine in 1040 ± 26.6 mg/L in in vitro in suspension cell cultures of C. roseus through the use of cyclodextrins and methyl jasmonate together with the exposure to UV light [99] was evidenced. Catharantine was obtained from in vitro cell cultures, but it has been very difficult to reach a stable accumulation with significant amounts of the vindoline metabolite [100].
On the other hand, the interaction between the cytokinins and ethylene in the accumulation of alkaloids in in vitro cultures has been investigated and a higher increase in the accumulation of ajmalicine in cells sub-cultivated in the free medium of 2,4-D was obtained [101].
The cells that were specialized and differentiated with the addition of precursors can produce more secondary metabolites than the in vivo cultures [102]. Although cells submitted to differentiation show slower growth than those that are not differentiated, some studies demonstrated that their morphology may have an influence; for example, it has been established that the cylindrical (i.e., elongated) cells of C. roseus contain a higher concentration of alkaloids than the round cells [103]. The inclusion of the tryptophan precursor may increase the content of catharantine [104] and it may also increase the number of elongated or cylindrical cells.
As far as we know, the way in which the content of catharantine is increased because of the tryptophan treatment and the release of other secondary metabolites on the same culture or the culture of cellular aggregates of C. roseus, remains to be well characterized [104].
Therefore, in vitro suspended cell cultures of C. roseus are considered as an excellent biotechnological tool for the in vitro and in vivo analyses of the biosynthetic pathway for the formation of anti-tumor alkaloids. With the aid of this tool, the relevant enzymes in the biosynthesis pathway were identified, but other studies are required to get into further detail at relevant enzymatic steps, even for the production of the anti-tumor drug taxol [105, 106]. For a comprehensive list of compounds isolated from these systems, also including the previously unidentified catindine and cavincidine compounds see [8].
ELICITATION OF ANTI-CANCER ALKALOIDS OF C. roseus IN in vitro CULTURES
Some new, recently discovered secondary metabolites of C. roseus have been obtained and their biological function established (Table 2).
Semi-synthesis and the organic synthesis of vincristine and vinblastine are highly costly and of low production yields, and their extraction from C. roseus is complicated, due to the low concentrations of these compounds within the plant. Therefore, alternative biotechnological strategies have been used to increase their levels as secondary metabolites, by adding biotic and abiotic inducers that stimulate the production of the metabolites in the alkaloids biosynthesis pathway (Table 3).
For example, the inclusion of pectinase increases 2.5 times the levels of tabersonine; the inclusion of chitin makes the levels of ajmalicine rise in 50 %, and the addition of jasmonic acid increases the levels of lochnericine and horthammericine, but not those of tabersonine [121].
Additionally, alkaloids have been detected in the pollen of C. roseus, mainly vincristine [122], which may provide a breakthrough for the pharmaceutical industry. Furthermore, phytochemical ingredients were identified, such as: total proteins, free amino acids, among others. The steroids and total pollen proteins were extracted by spectrophotometry, yielding 35.56 %. Free amino acids analysis detected 17 amino acids identified by the TLC method [122].
It has been demonstrated that the biosynthesis of the terpene indole alkaloids in C. roseus is tightly controlled at the cellular, tissue and organ levels [84], depending greatly on the phenology (the plant’s own biological state of development) and on the surrounding environment (biotic, abiotic and epigenetic factors).
On the other hand, it is important to remark that in feedback studies, it was found that high concentrations of the intermediaries or precursors of metabolites are able to generate greater accumulation of alkaloids in the biosynthesis pathway. But some experiments reveal that the presence of high concentrations of intermediary metabolites does not stimulate biosynthesis. In fact, the high content of metabolites, such as tryptamine and geraniol in culture media in vitro leads to cellular toxicity. This is attributed to the lack of differentiation of tissues, the direct exposure to metabolites or the high permeability of the membrane, but the most important aspect that is not considered is the lack of storage capacity of the synthesized tissue [84].
The production of alkaloids by in vitro cell culture may be considered as a differentiation process which is ruled by the environmental conditions (culture medium, phyto-hormones, light, temperature, precursors). The accumulation of alkaloids is a complex phenomenon, which is not totally disconnected to cell growth, but it is difficult to connect it with the appearance or disappearance of some metabolites or substances in the medium [84].
The production of alkaloids and the accumulation of biomass in vitro are directly influenced by the pH units of the medium; values with a range of 5.5 to 6.5 do not have much effect on the production of alkaloids. The value of 5.5 has been optimum for the production of serpentine [123].
It has been reported that the alkaloids produced in in vitro cultures of cells in suspension are stored in the vacuole and the storage capacity simultaneously changes when pH varies in the medium and the vacuole. The maximum and minimum units of pH were used by Asada and Shuler [124], to release the alkaloids within the cells in the culture medium. The optimum value (5.5-5.8) sometimes varies during the culture period and it influences the in vitro response on the production of terpene indole alkaloids [124].
METHODS OF QUALITATIVE AND QUANTITATIVE IDENTIFICATION OF THE CONTENT OF TERPENE INDOLE ALKALOIDS IN C. roseus
In the following are present some of the most common methods which make the quantification of indole alkaloids secondary metabolism and their products in C. roseus feasible. Vinblastine and vincristine have been isolated in a pure form by the use of several chromatographic techniques such as: vacuum liquid chromatography with a silica gel column: aluminum oxide (1:1) mixed with vacuum liquid chromatography (VLC); carbon column and purification by radial chromatography accelerated by centrifugation (Cromatotron). Semi-quantitatively procedures have been also implemented by using thin layer chromatography (TLC) methods [125]. TLC is highly sensitive to the detection of alkaloids; the ajmalicina is detected at 0.0007 % in a volume of 10 µL. The vincristine at 0.055 % in a volume of 10 µL while vinblastine and vindoline are not sensitive to this method because they are both found at concentrations of 0.05 % in a volume of 10 µL [126]. The chromogenic reagent that is chromatographically used in the detection of alkaloids is the cerium ammonium sulphate (CAS) that is known to react with the analyte to produce visible colors in the TLC plaque [15].
Other methods involve the analysis of the indole alkaloids by High Performance Liquid Chromatography (HPLC) [85]. The equipment for this method comprises a self-sampler able to analyze multiple samples. The separation of the
indole alkaloids is based on the reverse phase chromatography using C18 columns as the steady phase. The mobile phases usually consist of a mixture of buffer solutions such as n-heptanesulfonic acid, diammonium phosphate, or ammonium acetate supplemented with triethylamine and an organic phase (methanol or acetonitrile). The detection is carried out using a fixed wave UV detector or a fluorescent detector.
Additionally, the interaction of the growth hormones on the regulation of the indole alkaloids has also been extensively studied [13], as well as their extraction using super-critical fluids, which has been one of the most efficient quantification techniques used. Moreover, an understanding of the regulation of the metabolic flows is obtained with the metabolic flow analysis (MFA) [127], which requires the determination of the rates of the biosynthetic reaction. Nevertheless, a large number of measurements are required.
On the other hand, C. roseus has been phytochemically analyzed, and it has been found that it contains: carbohydrates, flavonoids, saponins, phenolic compounds, indole alkaloids, terpenoides [15], antocyanins (rosindin, 3-O-glucosides and 3-O-(6-O-p-coumaroyl) hirsutidin, malvidin and petunidin glucosides [16]. It lacks tannins, cardiac glycosides, steroids (catasterone, brasinolide) and monoterpene glucosides (loganin, secologanin, sweroside, dioxyl and dihydro-loganin) [8]. Two trisaccharide flavonoles of kaempferol and quercetin were also isolated [17, 60]; 15 glycosidic flavonoles were identified in seeds, stems, leaves and flowers [18], with other alkaloids been identified: 4’-deoxy-vinblastine, leurosine, pleurosine, leurocristine, leurosidine, vincoline, vincarodine, roseadine, vindolicine, rosicine, 5’-oxo-leurosine, N’b-oxide leurosidine and pericyclivine [6].
CONCLUSIONS
C. roseus has been a research icon because of the large number of phytochemical compounds, secondary metabolites and the therapeutic effects they produce. The secondary metabolites of C. roseus are terpene indole alkaloids with pharmacologic activity and with several applications in human medicine. The plant has a wide variety of properties: anti-cancer, anti-diabetes, anti-helminthic, anti-hypertensive, anti-diarrheic, anti-microbial, among others. The indole dimeric alkaloids: vinblastine and vincristine have become valuable drugs in cancer chemotherapy because of their potent anti-tumor activity against several types of leukemia and solid tumors. Remarkle examples are vinblastine, which is formed by catharantine and vindoline and catharantine, which is a member of the iboga family of the indole alkaloids.
Because of the use of plants and in vitro cell cultures, the biosynthesis pathway has been determined, but not completely clarified. Furthermore, a considerable number of enzymes have been characterized and their respective cloned genes defined, with the production of the alkaloids been found as highly regulated at the transcriptional level.
In order to increase the availability of alkaloids for therapeutic use, the production of biomass from in vitro cultures of calluses from leaves has arisen as biotechnological tool to increase the accumulation of alkaloids in C. roseus. The combination of different basal media as carbon sources, phytohormones and inducers of the biotic and abiotic type may provide useful ways for the rational technical development and the increase in production yields of several of these bioactive molecules in vitro. The in vitro cultures of calluses or cells in suspension could be used at a large industrial scale to obtain bioactive compounds that are of great importance in human health, and are envisaged as models to circumvent the limitations of other production systems.
REFERENCES
1. Taha HS, El Bahr MK, Seif-El-Nasr MM. In vitro studies on egyptian Catharanthus roseus (L.) G. Don.: 1-Calli production, direct shootlets regeneration and alkaloids determination. J Appl Sci Res. 2008;4(8):1017-22.
2. Villaseñor Ríos JL, Espinosa García FJ. Catálogo de malezas de México. México D. F.: Universidad Nacional Autónoma de México, Consejo Nacional Consultivo Fitosanitario, Fondo de Cultura Economica; 1998.
3. Acosta de la Luz L, Rodríguez C. Instructivo técnico para el cultivo de Catharanthus roseus (L.) G. Don. Vicaria. Rev Cubana Plant Med. 2002;7(2):96-9.
4. Fuentes V, Granda M. Conozca las plantas medicinales. La Habana: Editorial Científico-Técnica; 1997.
5. Granda M, Acosta L. Apuntes sobre el cultivo de plantas medicinales en Cuba. No. 4. Catharanthus roseus (L.) G. Don (vicaria). Rev Cult Trop. 1984;6(2):491-7.
6. Aslam J, Khan SH, Siddiqui ZH, Fatima Z, Maqsood M, Bhat MA, et al. Catharanthus roseus (L.) G. Don. An important drug: It’s applications and production. Pharmacie Globale. 2010;4(12):1-16.
7. Kuo YH, King ML. Antitumor drugs from the secondary metabolites of higher plants. In: Tringali C, editor. Bioactive compounds from natural sources: isolation, characterization and biological properties. London: Taylor & Francis; 2001. p. 189-282.
8. Van der Heijden R, Jacobs DI, Snoeijer W, Hallard D, Verpoorte R. The Catharanthus alkaloids: pharmacognosy and biotechnology. Curr Med Chem. 2004;11:607-28.
9. Karthikeyan B, Jaleel CA, Changxing Z, Joe MM, Srimannarayan J, Deiveekasundaram M. The effect of AM fungi and phosphorous level on the biomass yield and ajmalicine production in Catharanthus roseus. EurAsia J BioSci. 2008;2:26-33.
10. De Luca V, Cutler AJ. Subcellular localization of enzymes involved in indole alkaloid biosynthesis in Catharanthus roseus. Plant Physiol. 1987;85(4):1099-102.
11. Zhao J, Verpoorte R. Manipulating indole alkaloid production by Catharanthus roseus cell cultures in bioreactors: from biochemical processing to metabolic engineering. Phytochem Rev. 2007;6(2):435-57.
12. Soleimani F, Zarghami R, Ebrahimzadeh M. Effects of 2,4-D and kinetin concentrations on vinblastine and vincristine alkaloid contents in callus of periwinkle (Catharanthus roseus). Int J AgriSci. 2013;3(10):759-65.
13. Jaleel CA, Gopi R, Panneerselvam R. Growth and photosynthetic pigments responses of two varieties of Catharanthus roseus to triadimefon treatment. C R Biol. 2008;331(4):272-7.
14. Kisakurek MV, Hesse M. Chemotaxonomic studies of the Apocynaceae, Loganiaceae and Rubiaceae with reference to indole alkaloids. In: Phillipson JD, Zenk MH, editors. Indole and biogenetically related alkaloids. London: Academic Press; 1980. p. 11-26.
15. Ataei-Azimi A, Hashemloian BD, Ebrahimzade H, Majd A. High in vitro production of anti-canceric indole alkaloid from periwinkle (Catharanthus roseus) tissue culture. African J Biotechnol. 2008;7(16):2834-9.
16. Piovan A, Filippini R. Anthocyanins in Catharanthus roseus in vivo and in vitro: a review. Phytochem Rev. 2007;6(2):235-42.
17. Yadav PD, Bharadwaj NSP, Yedukondalu M, Methushala CH, Kumar AR. Phytochemical evaluation of Nyctanthes arbortristis, Nerium oleander and Catharanthus roseus. Ind J Res Pharm Biotechnol. 2013;1(3):333-8.
18. Ferreres F, Pereira DM, Valentao P, Andrade PB, Seabra RM, Sottomayor M. New phenolic compounds and antioxidant potential of Catharanthus roseus. J Agric Food Chem. 2008;56(21):9967-74.
19. Mallik J, Chowdhury HB, Al Farouq A, Das S. Pharmacological profile of Catharanthus roseus (apocynaceae) - a detailed review. Asian J Pharmaceut Res Dev. 2013;1(3):1-6.
20. Huxtable RJ. The pharmacology of extinction. J Ethnopharmacol. 1992; 37(1):1-11.
21. Leveque D, Jehl F. Molecular pharmacokinetics of Catharanthus (vinca) alkaloids. J Clin Pharmacol. 2007;47(5):579-88.
22. Casado P, Zuazua-Villar P, del Valle E, Martinez-Campa C, Lazo PS, Ramos S. Vincristine regulates the phosphorylation of the antiapoptotic protein HSP27 in breast cancer cells. Cancer Lett. 2007;247(2):273-82.
23. Martínez RA, Ceballos AA, Flores JA, Cuervo J, Gómez D. ¿Es la vinblastina útil en la leucemia aguda mieloblástica? Rev Hematol Mex. 2012;13(1):11-5.
24. Ferlay J, Shin HR, Bray F, Forman D, Mathers C, Parkin DM. GLOBOCAN 2008 v1.2, Cancer Incidence and mortality worldwide: IARC CancerBase No. 10 [Internet]. Lyon, France: International Agency for Research on Cancer. 2010 [cited 2014 Jan 15]. Available from: http://globocan.iarc.fr
25. Curado MP, Edwards B, Shin HR, Storm H, Ferlay J, Heanue M, et al. Cancer Incidence in Five Continents Volume IX. IARC Scientific Publications Nº 160. Lyon: International Agency for Cancer Research (WHO) and International Association for Cancer Registries; 2007.
26. IARC-OMS. World Cancer Report 2008. International Agency for Research on Cancer, Lyon, 2008.
27. American Cancer Society. Cancer Facts and Figures 2010. Atlanta: American Cancer Society. 2010.
28. Zelenetz AD, Abramson JS, Advani RH, Andreadis CB, Byrd JC, Czuczman MS, et al. NCCN Clinical Practice Guidelines in Oncology: non-Hodgkin’s lymphomas. Journal of the National Comprehensive Cancer Network : JNCCN. 2010;8(3):288-334.
29. American Cancer Society. Cancer Facts and Figures for Hispanic/latinos 2009-2011. Atlanta: American Cancer Society; 2009.
30. Rizo P, Sierra MI, Vázquez G, Cano M, Meneses A, Mohar A. Registro Hospitalario de Cáncer: Compendio de cáncer 2000-2004. Cancerología. 2007;(2):203-87.
31. Tirado-Gómez LL, Mohar Betancourt A. Epidemiología de las neoplasias hematológicas. Cancerología. 2007;(2):109-20.
32. Mousavi M, Ilkhani AR, Sharifi S, Mehrzad J, Eghdami A, Monajjemi M. DFT studies of nano anticancer on vinblastine and vincristine molecules. Int J Microbiol Res Rev. 2013;1(2):31-8.
33. Jordan MA, Wilson L. Microtubules as a target for anticancer drugs. Nat Rev Cancer. 2004;4(4):253-65.
34. Barrales-Cureño HJ, Ramírez Sepúlveda MF. A review on the production of taxoids anti-cancer of callus and cells of Taxus spp. in cultures in vitro. Rev Colomb Biotecnol. 2013;25(2):167-77.
35. Owellen RJ, Hartke CA, Hains FO. Pharmacokinetics and metabolism of vinblastine in humans. Cancer Res. 1977;37(8 Pt 1):2597-602.
36. Piccinini M, Tazartes O, Mezzatesta C, Ricotti E, Bedino S, Grosso F, et al. Proteasomes are a target of the anti-tumour drug vinblastine. Biochem J. 2001;356(Pt 3):835-41.
37. Vacca A, Lurlaro M, Ribatti D, Minischetti M, Nico B, Ria R, et al. Antiangiogenesis is produced by nontoxic doses of Vinblastine. Blood. 1999;94(12):4143-55.
38. Bates DJ, Salerni BL, Lowrey CH, Eastman A. Vinblastine sensitizes leukemia cells to cyclin-dependent kinase inhibitors, inducing acute cell cycle phase-independent apoptosis. Cancer Biol Ther. 2011;12(4):314-25.
39. Walts LR. The herbal encyclopedia. A practical guide to the many uses of herbs. New York: iUniverse, Inc. 2004.
40. Cuellar A, Lorincz C. Catharanthus roseus (L.) G. Don que crece en Cuba. Aislamiento y caracterización de vinblastina y leurosina, dos alcaloides con propiedades citostáticas. Rev Cubana Farm. 1975;9(3):183-99.
41. Singh SN, Vats P, Suri S, Shyam R, Kumria MM, Ranganathan S, et al. Effect of an antidiabetic extract of Catharanthus roseus on enzymic activities in streptozotocin induced diabetic rats. J Ethnopharmacol. 2001;76(3):269-77.
42. Kumari K, Gupta S. Phytopotential of Catharanthus roseus L.(G.) Don. var. “Rosea” and “Alba” against various pathogenic microbes in vitro. Int J Res Pure Appl Microbiol. 2013;3(3):77-82.
43. Amirjani MR. Effects of drought stress on the alkaloid contents and growth parameters of Catharanthus roseus. J Agric Biol Sci. 2013;8(11):745-50.
44. Siddiqui M, Ismail Z, Aisha AFA, Majid AMS. Cytotoxic activity of Catharanthus roseus (Apocynaceae) crude extracts and pure compounds against human colorectal carcinoma cell line. Int J Pharmacol. 2010;6(1):43-7.
45. Idrees M, Naeem M, Khan MMA. The superiority of cv ‘rosea’ over cv ‘alba’ of periwinkle (Catharanthus roseus L.) in alkaloid production and other physiological attributes. Turk J Biol. 2010;34:81-8.
46. Farnsworth NR. The pharmacognosy of the periwinkles: Vinca and Catharanthus. Lloydia. 1961;24(3):105-38.
47. Ponarulselvam S, Panneerselvam C, Murugan K, Aarthi N, Kalimuthu K, Thangamani S, et al. Dodder transmission of alder yellows phytoplasma to the experimental host Catharanthus roseus (periwinkle). Eur J Forest Pathol. 1997;27(6):347-50.
48. Alor MJ, Gómez R, Huerta E, Pat JM, González M, De la Cruz C. Nutrición y crecimiento en fase de vivero de Catharanthus roseus (L.) G. Don, Momordica charantia L. y Azadirachta indica A. Juss, en el Municipio Centro, Tabasco-México. Bol Latinoam Caribe Plant Med Arom. 2012;11(2):163-71.
49. Jaffe ES, Harris NL, Stein H, Isaacson PG. Classification of lymphoid neoplasms: the microscope as a tool for disease discovery. Blood. 2008;112(12):4384-99.
50. Cruz Ortíz H, Rodríguez-Martínez HA, Román Bassaure E, Valle Gay A. Coriocarcinoma gestacional con metástasis múltiples. Rev Fac Med UNAM. 2000;43(4):153-6.
51. Rocha JP, Leech MJ. Regulación transcripcional de genes involucrados en la producción de alcaloides indol-terpenoides en plántulas de Catharanthus roseus. Revista Colombiana Biotecnol. 2002;4(2):17-26.
52. Evans WC, Trease GE, Evans D. Pharmacognosy. 16th Ed. New York: Elsevier; 2009.
53. Taha HS, El-Bahr MK, Seif-El-Nasr MM. In vitro studies on Egyptain Catharanthus roseus L. G. Don. IV: Manipulation of some amino acids as precursors for enhanced of indole alkaloids production in suspension cultures. Aus J Basic Appl Sci. 2009;3:3137-44.
54. Hedhili S, Courdavault V, Giglioli-Guivarc’h N, Gantet P. Regulation of the terpene moiety biosynthesis of Catharanthus roseus terpene indole alkaloids. Phytochem Rev. 2007;6:341-51.
55. Vas A, Gulyas B. Eburnamine derivatives and the brain. Med Res Rev. 2005;25(6):737-57.
56. Gayatri CL, Chakravarthy R. Micro Propagation in Catharanthus roseus. Int J Innov Technol Explor Eng. 2013;2(5):194-6.
57. Kutney J, Mayer L, Schmidt B, inventors. Anhydrovinblastine for the treatment of cervical and lung cancer. International Patent WO 199839004. 1998 Mar 4.
58. Wong SK, Lim YY, Chan WCE. Botany, uses, phytochemistry and pharmacology of selected apocynaceae species: a review. Pharmacognosy Commu. 2013:3(3):2-11.
59. Roquebert J, Demichel P. Inhibition of the alpha 1 and alpha 2-adrenoceptor- mediated pressor response in pithed rats by raubasine, tetrahydroalstonine and akuammigine. Eur J Pharmacol. 1984;106(1):203-5.
60. Chung IM, Ali M, Chun SC, Lee SJ, San KY, Christie AMP, et al. New sesquiterpene glycosides from culture hairy roots of Catharanthus roseus. Chinese J Chem. 2007;25(11):1695-9.
61. Barrales-Cureño HJ, De la Rosa- Montoya, Villegas-Ortiz S. Hacia una genética celular del cáncer. Revista la Ciencia y el Hombre. 2012;25(2):1-6.
62. Moreno-Valenzuela OA, Galaz-Avalos RM, Minero-García Y, Loyola-Vargas VM. Effect of differentiation on the regulation of indole alkaloid production in Catharanthus roseus hairy roots. Plant Cell Rep. 1998;18(1):99-104.
63. Wang S, Zheng Z, Weng Y, Yu Y, Zhang D, Fan W, et al. Angiogenesis and anti-angiogenesis activity of Chinese medicinal herbal extracts. Life Sci. 2004;74(20):2467-78.
64. El-Merzabani MM, El-Aaser AA, Attia MA, El-Duweini AK, Ghazal AM. Screening system for Egyptian plants with potential anti-tumour activity. Planta Med. 1979;36(2):150-5.
65. Chattopadhyay SP, Das PK. Evaluation of Vinca rosea for the treatment of warts. Ind J Dermatol Venerol Leprol. 1990;56(2):107-8.
66. Cordell GA, Weiss SG, Farnsworth NR. Structure elucidation and chemistry of Catharanthus alkaloids. XXX. Isolation and structure elucidation of vincarodine. J Org Chem. 1974;39(4):431-4.
67. Agarwal S, Jacob S, Chettri N, Bisoyi S, Tazeen A, Vedamurthy AB, et al. Evaluation of in-vitro anthelminthic activity of Catharanthus roseus extract. Int J Pharm Sci Drug Res. 2011;3(3):211-3.
68. Hassan KA, Brenda AT, Patrick V, Patrick OE. In vivo antidiarrheal activity of the ethanolic leaf extract of Catharanthus roseus Linn. (Apocyanaceae) in Wistar rats. African J Pharm Pharmacol. 2011;5(15):1797-800.
69. Kumar A, Singhal KC, Sharma RA, Vyas GK, Kumar V. Analysis of antioxidant activity of Catharanthus roseus (L.) and its association with habitat temperature. Asian J Exp Biol Sci. 2012;3(4):706-13.
70. Rasool N, Rızwan K, Zubaır M, Naveed K, Ahmed V. Antioxidant potential of different extracts and fractions of Catharanthus roseus shoots. Int J Phytomed. 2011;3:108-14.
71. Muhammad LR, Muhammad N, Tanveer A, Baqir SN. Antimicrobial activity of different extracts of Catharanthas roseus. Clin. Exp Med J. 2009;3:81-5.
72. Patil PJ, Ghosh JS. Antimicrobial activity of Catharanthus roseus - A detailed study. Br J Pharm Toxicol. 2010;1(1):40-4.
73. Nayak BS, Pinto Pereira LM. Catharanthus roseus flower extract has wound-healing activity in Sprague Dawley rats. BMC Complement Altern Med. 2006;6:41-6.
74. Ramya S, Govindaraji V, Kannan KN, Jayakumararaj R. In vitro evaluation of antibacterial activity using crude extracts of Catharanthus roseus L. (G.) Don. Ethnobot Leaflets. 2008;12:1067-72.
75. El-Sayed A, Cordell GA. Catharanthus alkaloids. XXXIV. Catharanthamine, a new antitumor bisindole alkaloid from Catharanthus roseus. J Nat Prod. 1981;44(3):289-93.
76. Wong SK, Lim YY, Abdullah NR, Nordin FJ. Antiproliferative and phytochemical analyses of leaf extracts of ten Apocynaceae species. Pharmacognosy Res. 2011;3(2):100-6.
77. Jayanthi M, Sowbala N, Rajalakshmi G, Kanagavalli U, Sivakumar V. Study of anti hyperglycemic effect of Catharanthus roseus in alloxan induced diabetic rats. Int J Pharm Pharmaceut Sci. 2009;2(Suppl. 4):114-6.
78. Ghosh RK, Gupta I. Effect of Vinca rosea and Ficus racemososus on hyperglycemiain rats. Ind J Animal Health. 1980;19:145-8.
79. Chattopadhyay RR. A comparative evaluation of some blood sugar lowering agents of plant origin. J Ethnopharmacol. 1991;67(3):367-72.
80. Singh R, Kharb P, Rani K. Rapid micropropagation and callus induction of Catharanthus roseus in vitro using different explants. World J Agric Sci. 2011;7(6):699-704.
81. Tiong SH, Looi CY, Hazni H, Arya A, Paydar M, Wong WF, et al. Antidiabetic and antioxidant properties of alkaloids from Catharanthus roseus (L.) G. Don. Molecules. 2013;18(8):9770-84.
82. Shanks JV, Morgan J. Plant ‘hairy root’ culture. Curr Opin Biotechnol. 1999;10(2):151-5.
83. De Luca V, Laflamme P. The expanding universe of alkaloid biosynthesis. Curr Opin Plant Biol. 2001;4(3):225-33.
84. St-Pierre B, Vazquez-Flota FA, De Luca V. Multicellular compartmentation of Catharanthus roseus alkaloid biosynthesis predicts intercellular translocation of a pathway intermediate. Plant Cell. 1999;11(5):887-900.
85. Tikhomiroff C, Jolicoeur M. Screening of Catharanthus roseus secondary metabolites by high-performance liquid chromatography. J Chromatogr A. 2002;955(1):87-93.
86. Barrales-Cureño HJ, Soto RM, Ramos AC, Trejo LI, Martínez M, Ramírez ME, et al. Extracción y cuantificación de taxoides por HPLC en hojas in situ y en callos inducidos in vitro de Taxus globosa Schlecht. Spanish J Rural Dev. 2011;2:103-14.
87. Barrales-Cureño HJ, Soto HM, Ramos VA, Luna PG, Trejo TL, Martínez VM, et al. Inducción de callos in vitro de Taxus globosa a partir de acículas. Rev Latinoam Quím. 2009;Suplemento Especial: 89.
88. Fatemeh KA, Abdolreza B. The effect of hormonal composition, type of explant and light condition on callus production in Periwinkle (Catharanthus roseus L.). Plant Tissue Cult Biotechnol. 2013;23(1):107-13.
89. Haq R, Naz S, Aslam F, Manzoor F. Comparison of in vitro response of micropropagation and callogenesis of medicinal plant, Vinca rosea L. J Agric Res. 2013; 51(1):9-17.
90. Verma AK, Singh RR, Singh S. Improved alkaloid content in callus culture of Catharanthus roseus. Bot Serbica. 2012;36(2): 123-30.
91. Verma A, Laakso I, Seppanen-Laakso T, Huhtikangas A, Riekkola ML. A simplified procedure for indole alkaloid extraction from Catharanthus roseus combined with a semi-synthetic production process for vinblastine. Molecules. 2007;12(7):1307-15.
92. Negi RS. Fast in-vitro callus induction in Catharanthus roseus - A medicinally important plant used in cancer therapy. Res J Pharm Biol Chem Sci. 2011;2(4):597-603.
93. Kalidass Ch, Ramasamy MV, Arjunan D. Efecto de auxina y citoquinina en la producción de vincristina por cultivos in vitro de callos de Catharanthus roseus. (apocynaceae). Trop Subtrop Agroecosyst. 2010;12:283-8.
94. Ferderique H, Leslie M. Indole alkaloids biosynthesis. Plant Cell Environ. 1996;19:761.
95. Pandiangan D, Tilaar W, Nainggolan N. Morphological changes of cell in relation to increased catharanthine content of Catharanthus roseus cell aggregate cultures after tryptophan treatment. Int J Basic Appl Sci. 2013;13(1):45-51.
96. Hamada H, Tanaka T, Furuya T, Takahata H, Nemoto H. Hydroxylation of benzylic and allylic sites by plant cultured suspension cells. Tetrahedron Lett. 2001;42(5):909-11.
97. Ye M, Dai J, Guo H, Cui Y, Guo D. Glucosylation of cinobufagin by cultured suspension cells of Catharanthus roseus. Tetrahedron Lett 2002;43(47):8535-8.
98. Saifullah S, Kahn S. Callus induction and cell suspension culture production of Catharanthus roseus for Biotransformation studies of (-)- caryophyllene oxide. Pak J Bot. 2011;43(1):467-73.
99. Almagro L, Lopez Perez AJ, Pedreno MA. New method to enhance ajmalicine production in Catharanthus roseus cell cultures based on the use of cyclodextrins. Biotechnol Lett. 2011;33(2):381-5.
100. Shanks JV, Rijhwani SK, Morgan J, Vani S, Bhadra R, Ho CH. Quantification of metabolic fluxes for metabolic engineering of plant products. In: Fu TJ, Singh G, Curtis WR, editors. Plant cell and tissue culture for the production of food Ingredients. New York: Kluwer Academic/Plenum Publishers; 1999. p. 45-60.
101. Yahia A, Kevers C, Gaspar T, Chénieux JC, Rideau M, Crèche J. Cytokinins and ethylene stimulate indole alkaloid accumulation in cell suspension cultures of Catharanthus roseus by two distinct mechanisms. Plant Sci. 1998;133(1):9-15.
102. Moreno PRH, Van der Heijden R, Verpoorte R. Cell and tissue cultures of Catharanthus roseus: A literature survey II. Updating from 1998 to 1993. Plant Cell Tissue Organ Cult. 1995;42:1-25.
103. Kim SW, Jung KH, Kwak SS, Liu JR. Relationship between cell morphology and indole alkaloid production in suspension cultures of Catharanthus roseus, Plant Cell Rep. 1994;14(1):23-6.
104. Pandiangan D, Rompas DH, Aritonang HF, Esyanti RR, Marwani E. The influence of tryptophan on growth and catharanthine content in C. roseus callus culture. Jurnal Matematika dan Sains. 2006;11:111-5.
105. Barrales-Cureño HJ, Soto Hernández M. Bioquímica de los taxoides utilizados contra el cáncer. Rev Educ Bioquímica. 2011;30(1):12-20.
106. Barrales-Cureño HJ, Soto HM. Taxoids: secondary metabolites of the yew tree (Taxus spp.). Rev Chapingo. Serie ciencias forestales y del ambiente. 2012;18(2):207-21.
107. Wissing JB, Kornak B, Funke A, Riedel B. Phosphatidate Kinase. A novel enzyme in phospholipid metabolism (characterization of the enzyme from suspension-cultured Catharanthus roseus cells). Plant Physiol. 1994;105(3):903-9.
108. Marfori EC, Kajiyama S, Fukusaki E, Kobayashi A. Trichosetin, a novel tetramic acid antibiotic produced in dual culture of Trichoderma harzianum and Catharanthus roseus Callus. Z Naturforsch C. 2002;57(5-6):465-70.
109. Mitsuhashi N, Ohnishi M, Sekiguchi Y, Kwon YU, Chang YT, Chung SK, et al. Phytic acid synthesis and vacuolar accumulation in suspension-cultured cells of Catharanthus roseus induced by high concentration of inorganic phosphate and cations. Plant Physiol. 2005;138(3):1607-14.
110. Kargi F, Ganapathi B, Maricic K. Indole alkaloid formation by Catharanthus roseus cells in a biofilm reactor. Biotechnol Prog. 1990;6(4):243-8.
111. Chambault J, Volesky B, Kurz WG. Production of indole alkaloids by surface immobilized C. roseus cells. Biotechnol Bioeng. 1990;35(7):660-7.
112. Cau-uitz R, Miranda-Ham ML, Coello-Coello J, Chi B, Pacheco LM, Loyola-Vargas VM. Indole alkaloid production by transformed and non-transformed root cultures of Catharanthus roseus. In Vitro Cell Dev Biol. 1994;30(1):84-8.
113. Canel C, Lopes-Cardoso MI, Whitmer S, van der Fits L, Pasquali G, van der Heijden R, et al. Effects of over-expression of strictosidine synthase and tryptophan decarboxylase on alkaloid production by cell cultures of Catharanthus roseus. Planta. 1998;205(3):414-9.
114. Xu M, Dong J, Zhu M. Effect of nitric oxide on catharanthine production and growth of Catharanthus roseus suspension cells. Biotechnol Bioeng. 2005;89(3):367-71.
115. Srivastava NK, Srivastava AK. Influence of gibberellic acid 14CO2 metabolism, growth, and production of alkaloids in Catharanthus roseus. Photosynthetica 2007;45(1):156-60.
116. Ramani S, Jayabaskaran C. Enhanced catharanthine and vindoline production in suspension cultures of Catharanthus roseus by ultraviolet-B light. J Mol Signal. 2008;3:9-14.
117. Zhou ML, Shao JR, Tang YX. Production and metabolic engineering of terpenoid indole alkaloids in cell cultures of the medicinal plant Catharanthus roseus (L.) G. Don (Madagascar periwinkle). Biotechnol Appl Biochem. 2009;52(Pt 4):313-23.
118. Suttipanta N, Pattanaik S, Kulshrestha M, Patra B, Singh SK, Yuan L. The transcription factor CrWRKY1 positively regulates the terpenoid indole alkaloid biosynthesis in Catharanthus roseus. Plant Physiol. 2011;157(4):2081-93.
119. Zhou ML, Zhu XM, Shao JR, Wu YM, Tang YX. An protocol for genetic transformation of Catharanthus roseus by Agrobacterium rhizogenes A4. Appl Biochem Biotechnol. 2012;166(7):1674-84.
120. Guo ZG, Liu Y, Gong MZ, Chen W, Li WY. Regulation of vinblastine biosynthesis in cell suspension cultures of Catharanthus roseus. Plant Cell Tiss Organ Cult. 2013;112(1):43-54.
121. Shanks JV, Bhadra R, Morgan J, Rijhwani S, Vani S. Quantification of metabolites in the indole alkaloid pathways of Catharanthus roseus: implications for metabolic engineering. Biotechnol Bioeng. 1998;58(2-3):333-8.
122. Thaware JS. Screening of pollen of Catharanthus roseus L. – An anticancer plant. Int J Drug Discov Herbal Res. 2012;2(2): 403-7.
123. Doller G, Alfermann AW, Reinhard E. Production von indole alkaloid in callus kulturen von Catharanthus roseus. Planta Med. 1976;30:14-20.
124. Asada M, Shuler ML. Stimulation of ajmalicine production and extraction from Catharanthus roseus: effect of adsorption in situ, elicitors and alginate immobilization. Appl Microbial Biotechnol. 1989;30(5):475-81.
125. Aniszewski T. Alkaloids-secrets of life: Alkaloid chemistry, biological significance, applications and ecological role. Netherlands: Elsevier; 2007.
126. Magagula NL, Mohanlall V, et al. Optimizing production of alkaloids in cell suspension culture, callus culture, and Agrobacterium rhizogen transformed hairy root culture of Catharanthus roseus. Durban: Durban University of Technology; 2013.
127. Stephanopoulos GN, Aristidou AA, Nielsen J. Metabolic engineering. San Diego CA: Academic Press; 1998.
Received in February, 2014.
Accepted in January, 2015.
Hebert Jair Barrales-Cureño. Instituto de Genética. Colegio de Postgraduados Campus Montecillo. Km. 36.5 Carret. Mexico-Texcoco. Montecillo, Texcoco, State of Mexico 94420, Mexico. E-mail: barrales.hebert@colpos.mx
 All the contents of this journal, except where otherwise noted, is licensed under a Creative Commons Attribution License
All the contents of this journal, except where otherwise noted, is licensed under a Creative Commons Attribution License

{kind=link}
{kind=link}
{kind=link}
{kind=link}