










My SciELO
 Custom services
Custom servicesServices on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkBiotecnología Aplicada
On-line version ISSN 1027-2852
Biotecnol Apl vol.32 no.4 La Habana Oct.-Dec. 2015
TECHNIQUE
Adjusting the conditions of a system for the in vivo production of a nucleopolyhedrovirus of Spodoptera frugiperda (Lepidoptera: Noctuidae)
Ajuste de las condiciones de un sistema para la producción in vivo de un nucleopoliedrovirus de Spodoptera frugiperda (Lepidoptera: Noctuidae)
Carolina Ruiz1, Juliana Gómez-Valderrama1, Martha Chaparro1, Paula Sotelo2, Laura Villamizar1
1 Laboratorio de Control Biológico, Centro de Investigación Tibaitatá, Corpoica Km. 14 vía Mosquera, Colombia.
2 Pontificia Universidad Javeriana sede Bogotá, Colombia.
ABSTRACT
The Colombian Spodoptera frugiperda nucleopolyhedrovirus isolate SfNPV003, was used to develop a biopesticide with a strong potential for the control of this pest in the field, making it an alternative to chemical pesticides. Its marketing has, however, been limited due to the high in vivo virus production costs, which has a significant impact on product price. Hence, in this study, the conditions for the viral multiplication system were adjusted, in order to increase process yield and reduce costs. Initially, the effect of larval age and viral inoculum concentration on viral productivity was determined. Subsequently, the effect of the dietary regime for larvae development was established and finally, the effect of harvest time was evaluated on productivity and contaminant content. Sixteen-day-old larvae (fourth instar) were chosen considering this as an appropriate inoculation age, inoculum concentration was 1 × 107 occlusion bodies (OBs)/mL and a wheat germ-based maintenance diet was selected. Seven days post-inoculation (23 day-old larvae, fourth instar) was chosen as the optimum harvesting time of the infected larvae. Under these conditions the production capacity of viral particles was increased 95 times (from 9.5 × 1011 to 9.1 × 1013 OBs/month), allowing for a 20 % reduction in product price, thus improving its economic feasibility.
Keywords: baculovirus, corn armyworm, biological control, viral multiplication.
RESUMEN
El aislamiento colombiano de nucleopoliedrovirus de Spodoptera frugiperda SfNPV003 se emplea para desarrollar un bioplaguicida con alto potencial para el control biológico de esta plaga, como alternativa a los plaguicidas químicos. Sin embargo, su comercialización se ha visto limitada por los altos costos de la producción in vivo del virus, que repercuten de manera importante en el precio del producto. Por tales razones, se ajustaron las condiciones del sistema de multiplicación viral, para aumentar el rendimiento y reducir los costos. Se determinó el efecto de la edad larval y la concentración viral del inóculo sobre la productividad del virus. Posteriormente se estableció el efecto de la dieta de alimentación de las larvas, y se evaluó el efecto del tiempo de recolección de las larvas infectadas sobre el rendimiento del proceso y el contenido de contaminantes. El periodo de 16 días (cuarto instar larval) se estableció como la edad adecuada para la inoculación a una concentración de 1 × 107 cuerpos de inclusión (CIs)/mL y con la larvas bajo una dieta a base de germen de trigo para garantizar su mantenimiento. Se seleccionó el día 7 posinoculación (larvas de 23 días, cuarto instar larval) como el tiempo óptimo de recolección de las larvas infectadas. Bajo estas condiciones se incrementó en 95 veces la capacidad de producción de partículas virales (de 9.5 × 1011 a 9.1 × 1013 CIs/mes), lo cual permitió una reducción del 20 % en el precio de venta del producto, mejorando su factibilidad económica.
Palabras clave: baculovirus, cogollero del maíz, control biológico, multiplicación viral.
INTRODUCTION
The corn armyworm Spodoptera frugiperda (J. E. Smith, 1797) (Lepidoptera: Noctuidae) is the most important pest of the corn fields in Colombia, producing losses ranging from 13 to 60 % [1]. The unrestrained use of chemical insecticides for their control bring about high costs, environmental contamination and pest resistance [2, 3]. The use of baculoviruses, specifically the nucleopolyhedrovirus (NPV) therefore arises as a promising and environmentally safe alternative, since it is a specific arthropod virus that is not harmful to other organisms [4].
A large number of biopesticides based on baculoviruses have been developed worldwide for the control of different agriculturally important pests [5], although their use has been limited, and in some cases the products have disappeared from the market. One of the main problems is that these viruses are obligate pathogens, where an infection of a host is needed for their multiplication, for which reason the implementation of an in vivo production system is required. The in vivo system on the larvae of the insect is cumbersome and highly labor demanding, thereby increasing production costs, while showing contamination by other microorganisms and differences in viral productivity due to the lack of standardized procedures [6, 7].
A Colombian isolate of the nucleopolyhedrovirus of S. frugiperda codified as SfNPV003, was formulated as a wettable powder (WP), showing strong insecticide activity on pests under field conditions with over 70 % efficacy and a minimal impact on the beneficial entomofauna associated to the crop [8], as well as the efficient protection of the virus against the inactivation produced by UV rays [9]. The product is stable for 17 months under storage conditions at room temperature and it is compatible with commonly used agricultural chemicals for plant health problems in corn; it can be used in integrated crop management programs [10].
In spite of the high potential of this biological product, its scaling up and marketing depend on the efficient production system of its active principle. Production is currently carried out through an in vivo viral propagation system through the inoculation of eight-day-old S. frugiperda larvae (second instar), with the nucleopolyhedrovirus SfNPV003 at a 5 × 106 occlusion bodies (OBs)/mL concentration. Larvae maintenance is carried out using an artificial diet [11], and the dead individuals are harvested daily starting on the fifth day after the inoculation. The system, however, is highly labor-demanding in order to check each larva daily; the larvae collected have a wide variability in weight and size, and the high cost of the diet have negative effects on the production cost of the active ingredient.
Considering all the above mentioned aspects, the aim of this work was to improve yields and decrease costs of NPV viral production, through the adjustment of the mass production of the SfNPV003 virus, assessing the different factors, such as the diet, the age of the larvae at infection, the concentration of the viral inoculum and the harvest time of the infected larvae.
MATERIALS AND METHODS
Insect rearing
The larvae used for this study were obtained from a S. frugiperda colony established from the larvae collected from corn crops at the Espinal municipality (Tolima, Colombia). This colony was renewed with insects obtained in the field every six months; for this study we used the insects of the third generation (F3). These larvae remained under controlled feeding conditions with an artificial diet described by Greene et al. [11].
Virus
The virus used in this study was the semi-purified isolate of SfNPV003, which was amplified through the inoculation of larvae of S. frugiperda of the third stage using the droplet feeding method [12]. The dead larvae were crushed in sterile distilled water and homogenized. The mixture was filtered through a sheet of cloth to eliminate insect tissues debris, and the viral concentration was determined by counting in a Neubauer chamber [10].
Preparing a semi-synthetic alternative for the production of the SfNPV003 viral isolate
With the artificial diet used to rear the insect [11], four modified compositions were designed by modifying their components as described in table 1.
All modified diets included a lipid source considering that all insects require a dietary source of sterols [13]. The diet based on wheat germ included cholesterol and linseed oil and was based on the diet used for Spodoptera litura [14]. Considering the high cost of the cholesterol, the remaining three diets used only linseed oil, according to the diet described by Gómez et al. [15] for S. frugiperda.
To assess the effect of the feeding substrate on larval development, a bioassay with larvae of S. frugiperda of the third stage of the offspring was used; these were individualized, and from that time onward, they were fed four modified diets daily and remained under controlled conditions. During the setting-up period and every 4 days for 20 days, each one of the larvae was weighed. The experimental unit consisted of a plastic box with 10 flasks, each containing a S. frugiperda larva, and a food cube; there were three replicates per treatment. The experimental design was completely randomized.
To assess the effect of the diet on viral productivity, another bioassay was carried out, where a group of S. frugiperda larvae of the third stage of the offspring that underwent a 24 hour fast were given a viral suspension adjusted to 5 × 106 OBs/mL, using the droplet feeding method [12]. Each larva was later inoculated in a plastic flask; they were fed an artificial diet cube, according to the corresponding treatment, and they were incubated in a bioassay room. As of the fifth day of the inoculation, the larvae were checked daily, and each larva that died because of viral infection was weighed, crushed, and diluted in a known volume of sterile water. With the viral suspension obtained, they were counted in a Neubauer chamber, and with this result, and the weight of the larva, the productivity was calculated and expressed as OBs per g of larva tissue.
The experimental unit consisted of a plastic box with 10 glasses containing one S. frugiperda larva each and an artificial diet cube according to the treatment. The experimental design was completely randomized, with three replicates per treatment. One absolute control was used, which corresponded to the larva that had not been inoculated.
A nutritional analysis of the diets was also carried out, which included the following parameters: crude protein, dry matter (105 ºC), ashes, neutral dietary fiber, acid dietary fiber and ether extract. Finally, a cost analysis for each diet was made, using the composition of each one and comparing it to the standard artificial diet.
In order to select the composition producing the best conditions for viral propagation, we analyzed the average weights of each larva after their death, as well as viral productivity (OBs/g), larva mortality due to viral infection, and the cost of the diet per liter, using a decision matrix according to the methodology described by Dean and Nishry [16].
Assessment of the effect of larval age and viral concentration on productivity
In order to determine the influence of larval age and viral concentration on the productivity of the SfNPV003 nucleopolyhedrovirus, we carried out a bioassay following the above mentioned procedures, but inoculating S. frugiperda larvae of 4, 8, 12 and 16 days of age with suspension adjusted to 5 × 106 OBs/mL. A bioassay was later carried out with the individuals of the chosen larval age where 5 concentrations of the inoculum were assessed, namely: 1 × 105, 5 × 105, 1 × 106, 5 × 106 and 1 × 107 OBs/mL.
The experimental unit for both bioassays consisted of a plastic box with 15 glasses containing one S. frugiperda larva and a diet cube, each. The experimental design was completely randomized, with three replicates per treatment. There was an absolute control consisting of uninoculated larvae.
To select the concentration of the most appropriate inoculum and the age of the larvae, the variables evaluated were: the average weight of each larva after its death, viral productivity (OBs/g), and mortality of the larvae due to viral infection. The selection of the concentration of the inoculum was carried out by establishing the importance of each factor through the decision matrix described above.
Effect of harvest time on viral productivity
Harvest time of the larvae was estimated with the concentration of the selected inoculum, evaluating 7, 9 and 11 days after the inoculation and a treatment in which the larvae were collected at the time of death (D), that is to say, only those showing viral infection symptoms. To set up the bioassay we used the previously described methodology.
The experimental unit for the two bioassays consisted of a plastic box with 15 glasses that contained one S. frugiperda larva and a diet cube, each. The experimental design was completely randomized, with three replicates per treatment. There was an absolute witness, consisting of uninoculated larvae. The variables evaluated were the average weight of each larva after its death and viral productivity (OBs/g).
On the other hand, the amount of contaminants found in the collected larvae in each one of the treatments were evaluated, determining the bacterial, mold and yeast concentration by plate counting (c.f.u./ mL), following the methodology described by Santos et al. [10].
The yield of the productive process of the selected treatment was estimated on the basis of the installed capacity that enables the inoculation of 104 601 larvae/ month, using the following formula:
![]()
Where:
D: weight of the larva in grams
P: productivity expressed as occlusion bodies produced per gram of larval tissue
Statistical analysis
The normality of the data was verified with a Shapiro-Wilk test (95 %) and the homoscedasticity was determined through the Barlett’s test (95 %). Since both of the principles were met, an analysis of variance (ANOVA) was carried out and the means were compared using least significant differences tests and mean comparisons were made using Tukey, according to each case (95 %). The data that did not comply with the normality and homoscedasticity parameters were submitted to a non-parametric analysis of Kruskal-Wallis. The Statistix 8.1 software was used for all statistical analyses.
RESULTS AND DISCUSSION
Development of a semi-synthetic diet alternative for the production of the SfNPV003 viral isolate
Effect of the diet on larval development
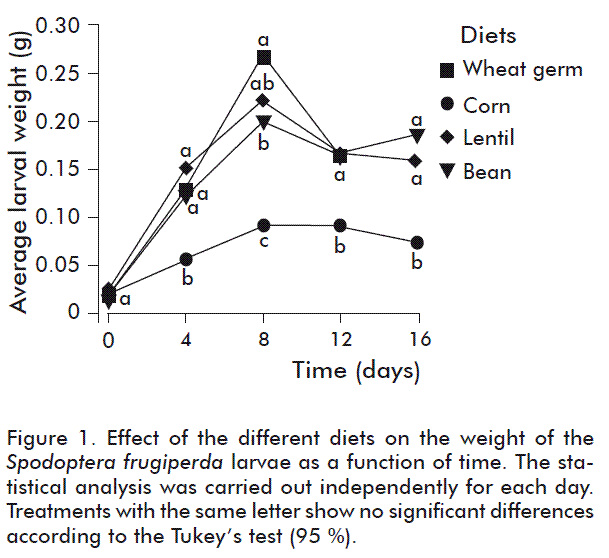
As of the time at which the different diets were supplied up until day 8, exponential weight gains of the larvae were observed in all treatments (Figure 1). The average weights of the larvae fed the diet based on wheat germ and the diet based on lentils, were similar and significantly higher (F = 18.56; d.f. = 3; p = 0.000) than the average weight obtained with the larvae fed the diet based on beans. The larvae fed the diet based on corn showed significantly lower weights on all evaluation dates, compare to the weights obtained with the other treatments (F = 18.56; d.f. = 3; p = 0.000) (Figure 1); this result suggests that the diet did not allow for an appropriate development of the S. frugiperda larvae.
As of day 12, and up until day 16, the larvae of all treatments started losing weight, possibly due to the start of the pre-pupa stage, in which the larvae stop eating and empty their intestine to enter the pupa stage [17].
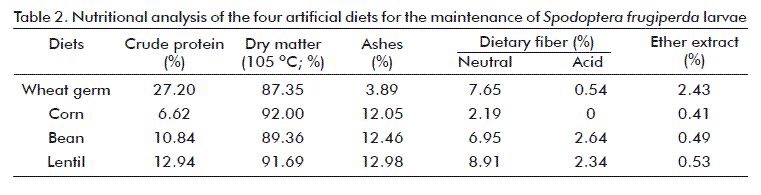
Nutritional composition analyses of the four diets used in this study are shown in table 2. According to these results, the best larval development was obtained with the larvae fed the diet based on wheat germ, which could be related to the differences in protein and ash content, acid dietary fiber and ether extract. Similar results were obtained by Busato et al. [18], who found a better larval development with S. frugiperda using diets with higher protein concentration, on evaluating two diets based on wheat germ.
It was found that the diet based on wheat germ had the highest percentage of crude protein, compared to the other diets (Table 2). Proteins are important sources of essential amino acids forming the body of the insect in their muscles, enzymes and membranes, while also being indispensable for survival, weight gains, cuticle formation, while it also participates in the immune system [19]. For example, Wiseman and Carpenter [20] studied the influence of the amount of protein in an artificial diet for Helicoverpa zea (Boddie) (Lepidoptera: Noctuidae) on larval development time, finding that this was significantly reduced on increasing the amount of protein, with an evident weight gain of the pupae, which is similar to the result of the current study.
Ash content was approximately 3 times lower in the diet based on wheat germ, indicating that this diet has less essential minerals; however, the proportion of these minerals needed for good larval development are trace amounts [21].
The percentage of neutral dietary fiber of the diets based on wheat germ, lentils and beans were higher than the diet based on corn (Table 2). Here is still another factor that may be associated to the low efficacy of this diet for the maintenance of the S. frugiperda larvae, since it has a lower amount of lignocellulosic and hemicellulosic compounds [22].
It is important to stress the result of the ether extract that was approximately five times higher in the diet based on wheat germ, compared to all the modified diets, showing that the cholesterol used in this diet generates a greater lipid contribution to the insect in relation to linseed oil, used in the modified diets. Lipids participate as a structural component of the cell membrane of the insect, while also being used as a source of energy and they are the predecessors of hormones [13, 21, 23, 24].
The diet based on cornmeal showed a nutritional unbalance because of its low nutrient content, which may have negatively affected the development and weight gains of the larvae, for which reason it was eliminated. Similar results were found by Murúa et al. [25], who evaluated four diets based on white bean meal, red corn meal (polenta), small-kerneled white corn (frangollo) and rice, finding that the larvae of S. frugiperda fed diets containing cornmeal showed supernumerary stages that were longer lasting, with 82 % mortality of the individuals during the formation of the pupal chamber.
Effect of the diet on viral productivity
Using the diets based on bean meal, lentils and wheat germ selected in the above bioassay, we evaluated their effect on viral productivity. The weights of the infected larvae as well as viral productivity were both affected by the diet used to feed the larvae. These variables were significantly higher when the larvae were kept under a wheat germ diet (F = 29.31; d.f. =2; p = 0.000) compared to the larvae fed the lentil diet and the bean diet (Table 3). These results suggest that the diet based on wheat germ had the best nutritional balance for the maintenance of the insect during the infection cycle. This is probably due to the appropriate availability of nutrients found in the substrate that are linked to the amount of cells/tissues available in the insect for the viral infection [26].
Concerning mortality, it was shown that the highest significant percentages were obtained with diets based on wheat germ and lentils with values of over 80 % (F = 13.9; d.f. = 2; p = 0.0056). This value is appropriate for this type of process, since most of the inoculated larvae are expected to be recovered dead because of the viral infection, reducing the loss of material [27]. With the diet based on bean meal, a significantly lower mortality was reached compared to other diets (F = 13.9; d.f. = 2; p = 0.0056), suggesting that this feed substrate alters the immunocompetency of the larvae due to the effect of nutrition, thus avoiding viral infection [19, 28].
In the decision matrix, the diet based on wheat germ had the highest final selection index (0.359) compared to the lentil meal diet (0.327) and the bean meal (0.315); that diet was therefore selected for larvae propagation, since it did not affect the development of the larvae, presenting the best nutritional content and producing the highest viral productivity at the lowest cost.
Effect of larval age and viral concentration on viral productivity
Significantly higher values were obtained with the individuals inoculated at 16 days compared to other inoculation ages evaluated for the weight variable (F = 142; d.f. = 3; p = 0.000) and for the viral productivity variable (F = 108; d.f. = 3; p = 0.000) (Table 4). As a consequence, we selected this age as the most appropriate one for the inoculation of the larvae for viral propagation. There is a constant relationship between the weight of the infected larvae at their death and the virus produced, since there is a greater availability of cells for viral replication [3, 29].
Afterwards, on evaluating 4 concentrations of the different inocula on this larval age, we found that there were no significant differences between any of the treatments evaluated (Table 5) (F = 0.34; d.f. = 3; p = 0.7936). In relation to mortality percentage, with the concentration of 1 × 107 OBs/mL we obtained a mortality of over 80 %, a value that is considered appropriate for this type of process, as explained above. In the decision matrix, the highest final selection index was obtained with the concentration of 1 × 107 OBs/mL (Table 5), for which reason this value was chosen as the inoculation concentration for the S. frugiperda larvae.
The effect of harvest time on viral productivity
After determining the diet, larval inoculation age and viral concentration of the inoculum, we evaluated the appropriate harvest time for the infected larvae to optimize productivity and minimize microbial contamination.
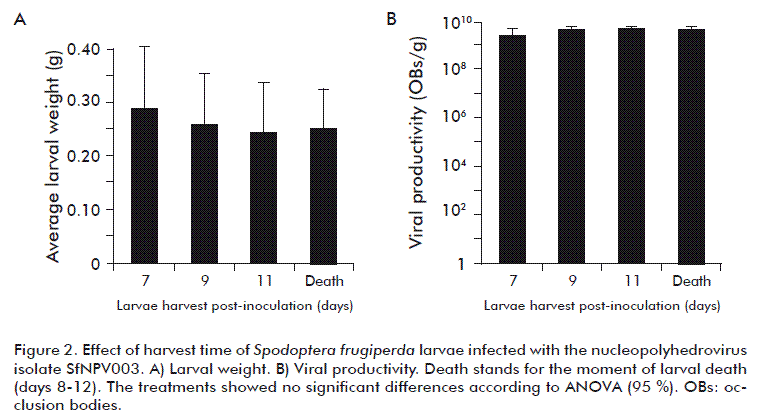
Figure 2 shows the results of the weight of the larvae (Figure 2A) and viral productivity (Figure 2B), corresponding to days 7, 9 and 11 after the inoculation and at the time of larval death (D), which took place between days 8 and 12 after the inoculation, where the dead individuals with symptoms of viral infection were collected. There were no significant differences between any of the treatments evaluated (F = 1.68; d.f. = 3; p = 0.2479), showing that the harvest time of the inoculated larvae did not have a significant effect on the amount of viruses recovered. However, the grams of larva collected at the end of the test was greater when the harvest was made 7 days after the inoculation with 12.51 g, which correspond to the harvest of 43 individuals, while when the harvest was carried out on days 9, 11 and the day of death the values obtained were of 7.83 g, 4.93 g and 3.14 g corresponding to 29, 21 and 13 individuals respectively. This is because when harvest time is increased after the inoculation, a larger number of individuals reached the pupal stage and were therefore not considered for the final analysis. Another important factor for the harvest of dead individuals with viral infection symptoms was the loss of the virus caused by the rupture of the integument, given the advanced state of degradation that was increased when the number of post-inoculation days increased [3, 30]. Similar results were found by Grzywacz et al. [6] who assessed the effect of incubation time on the viral productivity of the NPV of S. littoralis selected on day 7, which made it possible to obtain a higher productivity and facilitated the harvesting of the larvae.
One of the main difficulties in the in vivo production is the contamination with microorganisms from the collected material [31]; this is why in this study we evaluated the contaminant content found in the crushed larvae collected at each harvest time. The average results of this evaluation are shown in table 6. There were no significant differences in the amount of contaminating microorganisms present in the evaluated treatments (F = 1.23; d.f. = 3; p = 0.3603) (Table 6), but it was found that on increasing harvest time the number of contaminating microorganisms increased. Treatment D showed a numerically lower contamination since the harvesting of the individuals was made immediately after their death (Table 6).
This suggests that the larvae collected at their death have more microorganism contamination than the larvae collected before death or immediately afterwards. This may be due to the fact that the bodies of these insects become a substrate, promoting the growth of the saprophytic microorganisms, when large amounts of cellular material are exposed [6]. Furthermore, these saprophytic microorganisms may contaminate the nourishment substrate, the surface of the insect and the feces, increasing their concentration with time [30]. The elimination of these contaminants may generate an important increase in the cost of production, on having to implement corrective measures to decrease the concentration of microorganisms, as purification through filtration, centrifugation and the inclusion of antibiotics [30, 31]
Although the D treatment had a numerically lower amount of contaminants, the final volume of the virus collected was 75 % lower than that obtained on day 7 after the inoculation. Day 7 after the inoculation was chosen as the appropriate time to harvest the larvae infected with SfNPV003, on obtaining a larger volume of infected tissue, without any significant differences both in productivity and in the content of final contaminants, compared to the other treatments evaluated.
Taking into account the chosen treatment, with which individuals having an average weight per larva of 0.28 g and a productivity of 3.1 × 109 OBs/g of larval tissue were obtained, we calculated the monthly yield of the productive process with the adjustments made, and compared it with the previous process where we obtained larvae of 0.03 g with a productivity of 3.02 × 108 OBs/g of larval tissue. The yield of the process without any adjustment was estimated in a total production of 9.5 × 1011 OBs/month, which was increased 95 times on improving the process with the conditions chosen in the current study, reaching a yield of 9.1 × 1013 OBs/month.
CONCLUSIONS
With the selection of the diet (based on wheat germ), the age of the individuals at the time of the infection (16 days), the concentration of the viral inoculum (1 × 107 OBs/mL) and the harvest time allowing for a greater amount of the nucleopolyhedrovirus of S. frugiperda SfNPV003 and a lower contaminant content (day seven after the inoculation), we were able to increase the production capacity of viral particles per month 95 times. Here we used the same installed capacity, and it will produce a reduction of the sales price of the biopesticide in 20 %, improving its economic feasibility.
ACKNOWLEDGEMENTS
The authors would like to express their acknowledgement to the Administrative Department of Science and Technology COLCIENCIAS, to the Ministry of Agriculture and Rural Development of Colombia (MADR) and to the Colombian Corporation for Agricultural Research CORPOICA, for their financial support for this research.
REFERENCES
1. Instituto Colombiano Agropecuario ICA. RESOLUCION No. 000879. Bogotá: Ministerio de Agricultura y Desarrollo Rural. 2008.
2. Behle RW, Popham HJR. Laboratory and field evaluations of the efficacy of a fast-killing baculovirus isolate from Spodoptera frugiperda. J Invertebr Pathol. 2012;109:194-200.
3. Caballero P, Murillo R, Muñoz D, Williams T. El nucleopoliedrovirus de Spodoptera exigua (Lepidoptera: Noctuidae) como bioplaguicida: análisis de avances recientes en España. Rev Colomb Entomol. 2009;35:105-15.
4. Caballero P, Williams T, López-Ferber M. Los baculovirus y sus aplicaciones como bioinsecticidas en el control biológico de plagas. Madrid: España; 2001.
5. Copping L. The Manual of biocontrol agents, the biopesticide manual. 4th edition. Hampshire: The British Crop Protection Council; 2009.
6. Grzywacz D, Jones K, Moawad G, Cherry J. The in vivo production of Spodoptera littoralis nuclear polyhedrosis virus. J Virol Methods. 1998;71:115-22.
7. Sajjan DB, Hinchigeri SB. Increase in production of baculovirus occlusion bodies through cold incubation in silkworms. Int J Adv Res. 2015;3:5-17.
8. Gómez J, Guevara J, Cuartas P, Espinel C, Villamizar L. Microencapsulated Spodoptera frugiperda nucleopolyhedrovirus: insecticidal activity and effect on arthropod populations in maize. Biocontrol Sci Technol. 2013;23:829-46.
9. Villamizar L, Barrera G, Cotes AM, Martínez F. Eudragit S100 microparticles containing Spodoptera frugiperda nucleopolyehedrovirus: physicochemical characterization, photostability and in vitro virus release. J Microencapsul. 2010; 27:314-24.
10. Santos AM, Uribe LA, Ruiz JC, Tabima L, Gómez JA, Villamizar LF. Nucleopoliedrovirus de Spodoptera frugiperda SfNPV003: compatibilidad con agroquímicos y estabilidad en condiciones de almacenamiento. Corpoica Cienc Tecnol Agropecu. 2014;15:219-28.
11. Greene G, Leppla N, Dickerson W. Velvetbean caterpillar (Lepidoptera: Noctuidae) rearing procedure and artificial medium. J Econ Entomol. 1976;69:487-8.
12. Hughes P, Wood H. A synchronous peroral technique for the bioassay of insect viruses. J Invertebr. Pathol. 1981;37:154-9.
13. Cohen A. Insect diet: science and technology. Boca Raton: CRC Press LLC.; 2004.
14. Seth RK, Sharma VP. Growth, development, reproductive competence and adult behaviour of Spodoptera litura (Lepidoptera: Noctuidae) reared on different diets. Int Atom Energy Agency Tech Doc IAEA TECDOCs; 2002.
15. Gómez J, Guevara, Barrera G, Cotes AM, Villamizar L. Aislamiento, identificación y caracterización de nucleopoliedrovirus nativos de Spodoptera frugiperda en Colombia. Rev Fac Nac Agron. 2010;63:1-10.
16. Dean B, Nishry T. Scoring and profitability models for evaluating and selecting engineering products. J Oper Res Soc Am. 1965;13:550-69.
17. Gilbert L. Insect development morphogenesis, molting and metamorphosis. Oxford: Academic Press; 2009.
18. Busato GR, Garcia MS, Loeck AE, Zart M, Nunes AM, Bernardi O, et al. Adequacao de uma dieta artificial para os biótipos “milho” e “arroz” de Spodoptera frugiperda (Lepidoptera: Noctuidae). Bragantia. 2006;65:317-23.
19. Lee KP, Simpson SJ, Wilson K. Dietary protein-quality influences melanization and immune function in an insect. Funct Ecol. 2008;22:1052-61.
20. Wiseman B, Carpenter E. Growth inhibition of corn earworm (Lepidoptera: Noctuidae) larvae reared on resistant corn silk diets. J Econ Entomol. 1995;88:1037-43.
21. Lee KP, Simpson SJ, Raubenheimer D. A comparison of nutrient regulation between solitarious and gregarious phases of the specialist caterpillar, Spodoptera exempta (Walker). J Insect Physiol. 2004;50: 1171-80.
22. Pomeranz Y, Meloan C. Food analysis: Theory and practice; 3rd edition. New York: Chapman and Hall; 1994.
23. Karowe DN, Martin MM. The effects of quantity and quality of diet nitrogen on the growth, efficiency of food utilization, nitrogen budget, and metabolic rate of fifth-instar Spodoptera eridania larvae (Lepidoptera: Noctuidae). J Insect Physiol. 1989;35:699-708.
24. Popham HJR, Shelby KS. Uptake of dietary micronutrients from artificial diets by larval Heliothis virescens. J Insect Physiol. 2006;52:771-7.
25. Murúa MG, Virla EG, Defagó V. Evaluación de cuatro dietas artificiales para la cría de Spodoptera frugiperda (Lep.: Noctuidae) destinada a mantener poblaciones experimentales de himenópteros parasitoides. Bol San Veg Plagas. 2003;29:43-51.
26. Kalia V, Chaudhari S, Gujar GT. Optimization of production of nucleopolyhedrovirus of Helicoverpa armigera throughout larval stages. Phytoparasitica. 2001;29:23-8.
27. Grzywacz D, Rabindra RJ, Brown M, Jones KA, Parnell M. The Helicoverpa armigera NPV production manual. 2011 [cited 2015 Jun 15]. Available from: www.fao.org/docs/eims/upload/agrotech/2011/hanpvmanual-pt1.pdf
28. Adamo S. The impact of physiological state on immune function in insects. In: Rolff J, Reynolds S, Eds. Insect Infection and Immunity. Evolution, Ecology and Mechanisms. Oxford: Oxford University Press; 2009; p. 173-86.
29. Cherry J, Parnell M, Grzywacz D, Jones K. The optimization of in vivo nuclear polyhedrosis virus production in Spodoptera exempta (Walker) and Spodoptera exigua (Hubner). J Invertebr Pathol. 1997;70:50-8.
30. Caballero P, Williams T. Virus entomopatógenos. In: Control biológico de plagas agrícolas; Valencia: Editorial Phytoma; 2008. p. 121-35.
31. Ravensberg W. Progress in biological control. A roadmap to the successful development and commercialization of microbial pest control products for control of arthropods. Netherlands: Springer; 2011.
Received in July, 2015.
Accepted in December, 2015.
Carolina Ruiz. Laboratorio de Control Biológico, Centro de Investigación Tibaitatá, Corpoica Km. 14 vía Mosquera, Colombia. E-mail: jcruiz@corpoica.org.co.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}