










My SciELO
 Custom services
Custom servicesServices on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkBiotecnología Aplicada
On-line version ISSN 1027-2852
Biotecnol Apl vol.33 no.1 La Habana Jan.-Mar. 2016
RESEARCH
Micropropagation of Espeletiopsis rabanalensis S. Díaz & Rodr.-Cabeza
Micropropagación de Espeletiopsis rabanalensis S. Díaz & Rodr.-Cabeza
Eyda J Araque-Barrera, María de los A Bohórquez-Quintero, José C Pacheco-Maldonado
Laboratorio de Cultivo de Tejidos Vegetales Bioplasma, Universidad Pedagógica y Tecnológica de Colombia, Colombia.
ABSTRACT
Paramo ecosystems have been suffering alterations which represent a threat to the survival of species which inhabit them, among which we have frailejon, catalogued as key species to the restoration and preservation of biodiversity. Most frailejones are endangered species which require help for their reproduction through micropropagation, with the intention of massive multiplication. Bearing this in mind, a micropropagation protocol was established for Espeletiopsis rabanalensis, S. Díaz & Rodr.-Cabeza 2008, from embryos disinfected with Ca(OCl)2 at 4 % and cultured in MS/4 supplemented with GA3. Seedlings obtained were multiplied in MS supplemented with 0.5 mg/L IBA and 0.05 mg/L BA; 80 % of the shoots rooted in MS with 3 mg/L IBA and 75 % of the seedlings were viable after the hardening stage. The protocol established, along with previous works on Espeletiopsis muiska and Espeletia paipana effectively contribute to the recovery of endangered species, as well as the maintenance and protection of the water resource born in paramo ecosystems.
Keywords: frailejon, paramo, achene, embryo, micropropagation.
RESUMEN
Los ecosistemas de páramo han venido sufriendo alteraciones que representan una amenaza para la supervivencia de las especies que los habitan, entre las cuales se encuentran los frailejones, catalogadas como especies clave para restauración y conservación de la biodiversidad. La mayoría de frailejones son especies amenazadas que requieren ayuda para su reproducción a través de la micropropagación, con el fin de multiplicarlas masivamente. Teniendo en cuenta lo anterior, se estableció un protocolo de micropropagación para Espeletiopsis rabanalensis, S. Díaz & Rodr.-Cabeza, a partir de embriones desinfectados con Ca(OCl)2 al 4 % y cultivados en medio MS/4 suplementado con GA3. Las plántulas obtenidas se multiplicaron en MS con AIB (0.5 mg/L) y BA (0.05 mg/L), un 80 % de los brotes producidos enraizaron en MS con AIB (3 mg/L) y un 75 % de plántulas fueron viables después de la etapa de endurecimiento. El protocolo establecido, en conjunto con los trabajos en Espeletiopsis muiska y Espeletia paipana contribuye efectivamente a la recuperación de especies amenazadas, así como al mantenimiento y protección del recurso hídrico que nace en los ecosistemas paramunos.
Palabras clave: frailejón, páramo, aquenio, embrión, micropropagación.
INTRODUCTION
Paramos are high mountain ecosystems which constitute insular shelters and lack biological connectivity to similar ecosystems [1]. Both strategic location and extreme environmental conditions, as well as the ecosystemic goods and services they present, make paramo biomes of a great value in the regulation of water cycle and the retention of atmospheric carbon in the soil [2]. Nevertheless, paramos have been suffering severe alterations and transformations, and it is estimated that 66 % of these ecosystems have been modified [3, 4]. Such alterations have been causing physiological modifications in species and their life cycles, which pose a threat to their survival and, in a greater scale, for paramo ecosystems [5].
Frailejones (Espeletiinae) are endemic South American plants: Colombia (86 spp.), Venezuela (67 spp) and Ecuador (1 spp.) [6]. They show morphologic and physiologic adaptations to paramo conditions, where they thrive due to their stand and abundance. Due to their morphological characteristics and functionality, frailejones are a key species for the processes of restoration and preservation of high-mountain ecosystem biodiversity, since they hold great importance due to their trophic structure and in general. In fact, they enrich the goods and services provided by the paramo; therefore, their permanence is decisive for the maintenance of balance and ecological functions in paramo ecosystems [7]. Despite their importance in paramos, it is feasible that frailejones have been suffering alterations that influence their reproduction and survival. Currently there is no clear information available regarding causing agents, reason why it is hard to establish proper control strategies [5].
In Colombia, there are few studies regarding ecological restoration in high-Andean ecosystems [8] and although some projects have been presented to recover zones degraded due to mining use, the ones for paramounts are scarce [9]. Studies and restoration projects are aimed at restoring to degraded ecosystems its native functions and retrieving them their native natural conditions. For this reason, it becomes necessary to recuperate and/or maintain species such as E. rabanalensis, to maintain the ecosystem quality of paramos. Furthermore, there is a pressing need for the consolidation of strategies through developmental programs, which allow the application of biotechnologies, to enable faster, more precise and efficient processes to preserve, characterize, assess and productively take advantage of genetic resources [10]. Therefore, this work was aimed to establish a protocol for in vitro propagation, to support the development of replanting and preservation programs for E. rabanalensis in the Paramo Rabanal (Ventaquemada- Samacá, Boyacá), a paramo ecosystem that has been negatively affected by open pit coal mining.
MATERIALS AND METHODS
Study methodology
This research was carried out at BIOPLASMA-UPTC (Laboratorio de Cultivo de Tejidos Vegetales, Universidad Pedagógica y Tecnológica de Colombia). The methodology developed was based on the work by Araque et al. [11] and Bohórquez et al. [12] regarding Espeletia paipana.
Starting vegetal material, superficial disinfection and in vitro establishment
Adult plants of E. rabanalensis S. Díaz & Rodr.-Cabeza used as seed source are located in the Paramo Rabanal (Ventaquemada-Samacá, Boyacá, Colombia, at 3412 m of altitude). Between April and May, 2014, flower heads of E. rabanalensis were collected and mature achenes were extracted from them at the laboratory. From these achenes, embryos were extracted and superficially disinfected in sterile muslin bags, in batches of 30 units. In laminar flow chamber, bags containing the embryos were submerged for three min in 50 mL of sterile distilled water with 0.1 mL of Tween 20; then, they were rinsed three consecutive times with sterile distilled water. For superficial asepsis, the efficiency of calcium hypochlorite (Ca(OCl)2) was tested at 1 % (w/v; T1), 2 % (T2) and 4 % (T3), for five min and at 4 % (T4) for 7 min, as well as the effect of sodium hypochlorite (NaOCl) at 0.21 % (w/v) for 5 min (T5) and 7 min (T6). Finally, the bags containing the embryos were rinsed five consecutive times using sterile distilled water. Each asepsis treatment was applied to 924 embryos, for a grand total of 5543. Embryos of each treatment were cultured for 45 days in the following germination media: MS (Murashige and Skoog) [13] without growth regulators (1392 embryos), MS with salts diluted at ½ (MS/2) (1389 embryos), diluted at ¼ (MS/4) (1394 embryos) and finally MS/4 supplemented with 1 mg/L gibberellic acid (GA3;1368 embryos).
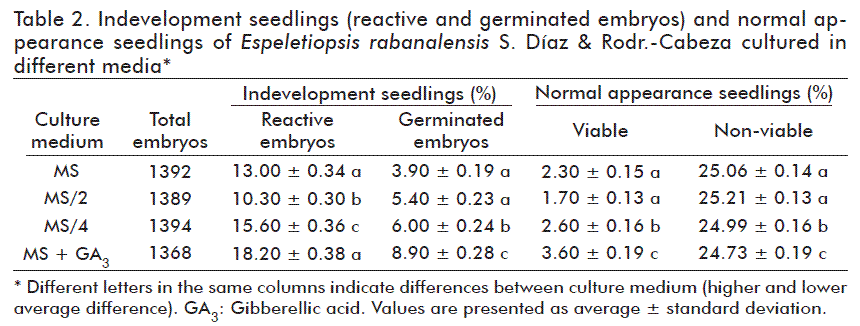
By the end of the germination stage (establishment stage), the percentages of aseptic embryos, fungus or bacteria-contaminated, non-viable, in-development seedlings (reactive and germinated embryos) and normal appearance seedlings were quantified.
Axillary shoot multiplication
Multiplication stage was performed using 1-2 cm long rootless seedlings developed in the previous stage. Explants were grown for six consecutive sub-cultures (30 days each) in MS without growth regulators and MS supplemented with 0.3 mg/L Indol-3-butyric acid (IBA) plus 0.03 mg/L 6-Benzylaminopurine (BA), 0.5 mg/L IBA plus 0.05 mg/L BA, and 1.0 mg/L IBA plus 0.1 mg/L BA; 30 explants (5 per a 100-mL glass jar with 15 mL of medium) were cultured in each medium. Taking into account the total amount of individualized shoots per sub-culture, the percentage of individualized shoot per treatment and the proliferation rate (PT) were calculated by using the relationship PT = (Bi – B0)/B0, with B0 and Bi representing the initial and individualized shoots, respectively. Individualized shoots was the expression used for shoots with an easily observable portion of stem (minimal length of 0.2 cm). Based on the results obtained, it was possible to determine the most appropriate hormonal combination for shoot multiplication, which was later used to support proliferation chains.
Shoot rooting and plantlet acclimatization
Individualized shoots obtained during the multiplication stage were selected for rooting and they were grown in MS without growth regulators and MS supplemented with 3, 5 or 7 mg/L IBA for 60 days with a subculture in fresh medium after 30 days. In each of the treatments performed, 30 explants were cultured and the percentage of rooted shoots was quantified, along with the number and length of roots per shoot and d the percentage of shoots which formed basal callus.
Rooted shoots were transferred to 200-mL glass jars, closed off with extensible paper which was progressively pierced to 80 % of its total surface. The jars were filled with 30 g of sterile washed sand and 15 mL of a solution containing 200 mg/L KNO3, 800 mg/L Ca(NO3)2 •4H2O, 200 mg/L KHPO4, 200 mg/L MgSO4•7H2O and 100 mg/L FeSO4. Jars were kept in an incubation room for 40 days at 23 ºC with continuous illumination. Subsequently, under greenhouse conditions, the plantlets were transferred to 250 cc plastic pots, each containing sterile substratum composed by ground mulch, sand and soil, 3:2:1; the plantlets were then watered manually once a day. After 60-day acclimatization, the percentage of viable plantlets was registered.
Culture conditions
Before adding agar (6.5 g/L) pH in every medium was adjusted to 5.7 with 1 N KOH or 1 N HCL; conse-quently, the media were sterilized in autoclave at 15 psi and 121 ºC for 20 min. Cultures were incubated at 24 ± 1 °C with continuous fluorescent illumination of 70-80 μmol/m2•s.
Data analysis
Data obtained were processed using PASW Statistics 18 Software, version 18.0.0 (jul-30-2009), and Excel 2013. Then the data were analyzed using the Chi-squared test goodness of fit with a confidence interval of 95 %. Once the differences between the average quantities were determined, they were grouped by higher and lower average difference in order to identify homogeneous subset.
RESULTS
Vegetal material, superficial disinfection and in vitro establishment
Up to 5544 embryos, 1-1.5 mm long, were retrieved from the achenes (Figure 1A) extracted from collected flower heads. These embryos were apt for in vitro culture, characterized by having consistent tissue, swollen aspect and whitish coloration (Figure 1B).
According to the Chi-square test, statistically significant differences were found between treatments (p = 0.0000282) treatments assayed for superficial asepsis (Table 1); nevertheless, similar behaviors were detected for treatments T1 and T2, T3, and for treatments T4, T5, and T6. The most effective treatments were T4 and T6, with 94.0 (868) and 85.0 % (785) aseptic embryos, followed by T3, T5, T2 and T1 with 74.0 (684), 73.0 (674), 47.0 (434) and 45.0 % (415) aseptic embryos, respectively. Similarly, 26 % (1441 out of the 5543 embryos treated) showed microbial contamination, 16 % (887) bacterial contamination and 10 % (554) fungal contamination (Table 1).
After 45 days of culture in germination media, 85.72 % (4753 embryos) did not show reactivity and remained with the same appearance they had at the start of culture. Developing seedlings showed greenish coloration or a slight elongation (14.28 %; 790), 42.27 % (334) of them germinating and 57.73 % (193) became brownish-grey and died, for a final 42.21 % (141) of seedlings with normal appearance after 90 days of culture. This accounted for 2.5 % of the total embryos cultured.
The highest percentage of seedlings under development (reactive embryos, 18.2 %; germinated embryos, 8.92 %; normal appearance seedlings, 3.58 %) were obtained in MS/4 supplemented with 1 mg/L GA3. The second best results were obtained in in MS/4 medium, with 15.6 % reactive embryos, 5.6 % germinated embryos and 2.6 % viable seedlings. Seedling growth and development with vigorous appearance and leaves in rosette arrangement were observed in these two culture media. Noteworthy, there were no statistically significant differences in the number of reactive embryos (Chi-square test; (p = 0.48966671) or germinated embryos (p = 0.52912812) as a function of the culture medium assayed. Nevertheless, there were variations between treatments the in reactivation and germination responses of embryos, which made possible distinguishing three groups according to percentage differences (Table 2). For the case of viable, normal appearance seedlings after 90 days, it was not possible to apply the Chi-square test, since the expected average values were lower than 5. For this reason, the data were processed and analyzed considering the unviability as a function of the assessed treatments, estimating that the culture media did not affect seedling unviability (p = 0.9993).
At the beginning of the germination process, embryos suffered a slight elongation and turned green-yellowish (Figure 1 C), cotyledons then separated, expanded and turned green (Figures 1 D-E), hypocotyls elongated and the development of the radicle began (Figures 1 F-G); the activity in the apical bud was evident and the first leaves developed (Figures 1 G-H); the stem reached 1-3 mm and new dark-green pubescent leaves developed in a helicoid arranged (Figures 1 H-K), reaching up to 3-cm height.
Multiplication of axillary shoots
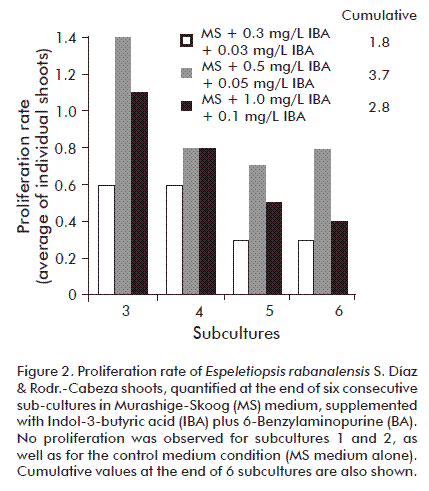
The new shoots were individualized from the third sub-culture on. Proliferation rates for the assayed treatments showed statistically significant differences for sub-cultures 3 (p = 0.000266129), 4 (p = 2.57775 × 10-15), 5 (p = 2.57775 × 10-15) and 6 (p = 2.57775 × 10-82). The treatment with the highest percentage of individualized shoots per culture (subculture 3: 33.49; subculture 4: 37.22; subculture 5: 42.26; and subc6: 50.06) was the one performed in MS supplemented with 0.5 mg/L IBA plus 0.05 mg/L BA (71, 131, 221 and 404 shoots in sub-cultures 3, 4, 5 and 6, respectively); by the end of the sub-culture 6, there was an accumulated rate of proliferation of 1:4 individualized shoots (Figure 2). Besides, in this treatment, individualized explants reached up to 4 cm height and showed rosette growth (Figures 1 L-M), 10-15 succulent, pubescent, 2-5 cm long, dark-green leaves with continuous edges (Figures 1 N-O).
Shoot rooting and plantlets acclimatization
After 25 days of culture in rooting media, most shoots showed radical primordia (Figure 1 P), root development (Figure 1 Q) was detected after 40 days in all assayed media, except the controls, reaching up to 90 % of rooted shoots in MS supplemented with 7 mg/L IBA and 86.7 % in MS plus 5 mg/L IBA (35.1 and 33.8 % respectively, from total number of rooted shoots). There were statistically significant differences (Chi-square test; p = 0.0358) between treatments, particularly with 5 mg/L IBA which induced the best average of root number and length. Keeping in mind the minimum length of roots, there were mild statistical significances (p = 0.0537; Chi = 7.6515) and considerable statistical significances for the maximum length (p = 0.0156; Chi = 10.3718). Nonetheless, similar minimum length roots were obtained in treatments with MS medium supplemented with either 3 or 7 mg/L IBA and similar maximum length roots when the plantlets were cultured in MS plus 3 or 5 mg/L IBA (Table 3).
Results quantified in MS plus 5 mg/L or 7 mg/L IBA showed the best results of shoot rooting, and also the highest amount of developed shoots with basal callus (statistically significant differences were found between treatments; p = 0.000547984). These two groups also differentiated from the others in the for-mation of basal calluses. Thus, the most appropriate medium for rooting of E. rabanalensis shoots was MS supplemented with 3 mg/L IBA (Table 3), this medium supporting the development of numerous, nor-mal-looking roots of considerable length, whitish and showing multiple absorbent hairs (Figure 1 R and S).
In plantlets rooted in MS supplemented with 3, 5 or 7 mg/L IBA, viability was 75, 15 and 10 %, respectively. Plantlets showed vigorous appearance, pubescent leaves, growth and development of shoots and roots with multiple absorbent hairs (Figures 1 T-U). Besides, shoots which formed basal callus were not viable during hardening, despite they developed roots.
DISCUSSION
When explants coming from adult plants are cultured in vitro, establishing completely aseptic cultures is usually very difficult due to the action of bacteria and fungi already present in those plants [14-16]. Nevertheless, in the case of E. rabanalensis, the application of sodium hypochlorite and calcium hypochlorite in adequate amounts is an effective alternative for the efficient superficial disinfection of sexual embryos. This last effect results from the germicide action of chlorine, which is less phytotoxic than other disinfectants and destroys microorganisms through its action on specific cellular components [17]. The use of calcium hypochlorite favored the recovery of a high percentage of reactive aseptic embryos with a smaller alteration and damage to their tissues, similarly to reports by Sánchez et al. [18] in Gyrocarpus americanus and Araque et al. [11] and Bohórquez et al. [12] in E. paipana. Furthermore, according to Sánchez and Salaverría [19], calcium hypochlorite might be part of some processes of vegetal metabolism, since Ca2+ is essential for several ones; calcium hypochlorite may also provide soluble calcium to the tissue, thereby neutralizing organic acids and the production of phenols and polyphenols, substances which contribute to the oxidation of the explants during superficial asepsis.
Although there were a high percentage of superficially aseptic embryos in the disinfection treatments here assayed, it was also observed that when cultured in vitro, the number of reactive and germinative embryos was low, a reason why the amount of viable seedlings (2.5 %) 90 days after culture was notoriously low. This response was similar to the one observed in E. paipana [12]. Nevertheless, this action is not applicable to E. rabanalensis since the inter-annual flowering rates are notoriously low and the reproductive sequence is asynchronous and irregular [20, 21], despite the quality of this action is essential to establish in vitro cultures from seeds. Therefore, the number of available seeds is limited and plants further show monoecious flower heads which vary in size and number of flowers [22]. They also present a strong sporophytic self-incompatibility mechanism and a tendency to hybridation [22, 23]. Likewise, García et al. [24] have pointed out that for Dendrocalamus strictus, the availability of seeds is affected by the long and erratic flowering cycle of the species, which makes its sexual propagation difficult; besides, it is also important to note that, in tropical settings, seeds lose viability after 2 or 3 months [25].
Noteworthy, in addition to the low germination capacity of the embryos, it is not possible to take buds, organs or tissues from E. rabanalensis to establish in vitro cultures. Instead, is relevant to use of zygotic embryos as initial explants for the establishment of in vitro cultures, since it allows maintaining, at least in part, the natural genetic diversity of the plant, a favorable aspect in the processes of repopulation of natural areas and for in situ and ex situ preservation [26]. Guerra et al. [27] pointed out that micropropagation systems are an important tool for the improvement and massive clonal propagation of selected genotypes, and that the use of embryos can be justified when conventional propagation methods were inefficient in the species. That is the case of frailejones and orchids, which bud multiplication depends on seed germination for the initial production of seedlings. It is worth to say that not all the seeds in a flower head, or a capsule for the case of orchids, are fully formed or fertile, and only those with a viable embryo actually germinate [28]. In other cases, the embryo is too small compared to the seed coat that the volume of the seed is formed for up to 96 % of air [29], thereby humidity not reaching the embryo, as in the case of Laelia speciosa [30]. Therefore, the obtainment of seedlings through in vitro seed germination is an appropriate tool when it is used to propagate hybrid plants or plants with very particular properties as E. rabanalensis.
Regarding the culture media assayed for embryo germination, those with mineral salts diluted at ¼ (MS/4) and supplemented with GA3 produced the best results for in vitro cultures establishment. These results matched the ones obtained by Mayo et al. [31], Brar et al. [32] and Bohórquez et al. [12] who established a high average of seed germination in low-salt media. Besides, Vidales [33] indicated that, in order to carry out micropropagation in some species, it should be advisable to use GA3 during the in vitro and ex vitro establishment phase, since phyto-regulator causes a positive effect by improving the germinative energy of the species mainly at the beginning of the germination process and during explant development. In this regard, Gamborg et al. [34] and Roca and Mroginsky [35] reported that success in the establishment of in vitro cultures depends on the medium selection, including chemical composition and physical form. Nevertheless, it is difficult to find a completely efficient medium for plant production from seeds, a particularly difficult task for the propagation of wild species.
During the shoot multiplication phase, explants coming from germinated embryos which showed contamination by bacteria were not viable; they suffered necrosis and died after the first or second sub-culture. In contrast, explants coming from germinated embryos which did not show contamination formed and developed new axillary buds, capable of responding to hormonal stimuli from the culture medium and become individualized shoots. Statistical resemblance between multiplication responses in the different treatments indicates that the range of usable hexogen hormonal balance is not so narrow, indicating that both high and low relative concentrations of assayed IBA and BA are effective to induce favorable response on shoot proliferation of E. rabanalensis. Regarding concentrations, Hugues [36] established that the levels of auxin and cytokinin required for morphogenesis in explants from different taxa and different genotypes were highly variable. Hence, such levels must be determined for each species or variety. Likewise, the positive effect of the auxin-cytokinin combination on cellular multiplication and the relevance of an adequate auxin/cytokinin balance to increase proliferation rates have been corroborated for micropropagation of Espeletiopsis muiska [37] and E. paipana [12], where a hormonal balance favorable towards cytokinin was determined.
On the other hand, as shown in figure 2, there was a growing tendency in the proliferation rate of individualized shoots produced in the MS médium supplemented with 0.5 mg/L IBA plus 0.05 mg/L BA, with bud formation pattern and shoot development consistent and repetitive in such a way that individualized shoots also formed axillary buds and developed new shoots. This guaranteed the continuous maintenance of proliferative chains and the production of shoots for in vitro rooting of E. rabanalensis.
Regarding the media assayed for the rooting process, it was observed that none of the shoots cultured in the control medium developed roots; therefore, it was evident that IBA in the assayed rooting media stimulated rhizogenic development of shoots on E. rabanalensis. This observation matches the statement made by Murashige [38], who determined that high relative concentrations of auxin favor root development. Data regarding E. rabanalensis shoot rooting showed statistically significant differences between treatments; nevertheless, it is necessary to bear in mind that the excess of auxin in the medium induced the formation of basal callus, which according to Rios et al. [39] and Bohórquez et al. [12] makes seedlings unviable during the processes of acclimatization and transference to greenhouse or open field conditions. Data registered in table 3 indicate that the number and length of the roots increases with the incremented concentration of auxin, similar to the process reported for E. muiska [37] and E. paipana [12], as long as concentration does not exceed 5 mg/L. It may be observed that the average values of the variables studied in E. rabanalensis decreased in presence of 7 mg/L IBA. Moreover, Németh [40] and Bennett et al. [41] indicated that in vitro rooting of the obtained shoots is an essential step which directly affects the success of in vivo plant regeneration.
Observations performed on rooted shoots indicate that both the development of a proper radical system and the development of functional roots, which facilitate nutrient absorption from substratum used during acclimatization, increment the percentage of viable seedlings in ex vitro conditions. When properly done, the acclimatization phase facilitates the formation and development of new functional structures which favor the viability of seedlings produced in vitro. That is the case of newborn leaves which develop cuticle, produce epicuticular wax and form functional stomata which provides an adequate transpiration rate. At the same time, it must be taken into account that plants generated by in in vitro culture behave as heterotrophic or mixotrophic organisms, a reason why during the acclimatization phase they are forced to become autotrophs, what changes their morphology and makes them susceptible to different types of stress.
The substrate used for acclimatization (ground mulch, sand and dirt; ratio 3:2:1) resulted appropriate for growth and development of E. rabanalensis plantlets. According to Valencia and Ramirez [42], this mixture provides high porosity, high water retention, good ventilation, low pH and a fine texture. In fact, the nutritional requirements of E. rabanalensis are rather particular, considering that it is a paramo species. For instance, Muñoz [43] has pointed out that it is important to keep the natural conditions on the soil in which the species are intended to be spread and later develop, not always being necessary to enrich the soil with fertilizer or any other corrective elements. Results obtained in E. rabanalensis matched those reported for E. muiska [37] and E. paipana [12], emphasizing that the initial control of temperature, humidity and illumination is essential during the first stage of acclimatization. Later on it was shown that that seedlings transferred directly from the culture room to greenhouse conditions did not survive, even though the mixture used as a substrate was adequate to establish ex vitro plantlets of frailejon.
Hence, our results contribute to the preservation not only of E. rabanalensis, a key species, but of the paramos itself as the fragile ecosystem as it is. Moreover, micropropagation of E. rabanalensis for replanting, relocation and preservation of high-mountain ecosystems is an affordable and effective strategy for the restoration and/or preservation of altered and degraded areas, further contributing to the protection of water resources. According to Sturm and Rangel [2, 44], the implementation of such strategy can be efficient for ecosystems where regeneration processes are slow due to low net primary productivity, low temperatures, drastic day-to-night temperature oscillations and acid soils, as it is the case of the tropical high-mountain ecosystem.
In summary, the establishment of an in vitro propagation protocol for E. rabanalensis contributes to the knowledge and preservation of frailejones, facilitating the production of plants from embryos superficially disinfected with calcium hypochlorite (4 %) and cultured in MS/4 medium further supplemented with 1 mg/L GA3. The in vitro vegetal material can be multiplied through proliferative chains in MS with 0.5 mg/L IBA plus 0.05 mg/L BA; up to 80 % of shoots can produced roots in MS medium supplemented with 3 mg/L IBA and 75 % viable plantlets can be retrieved by the end of the acclimation stage. These also support further studies to improve and maintain the native, introduced or endangered species usable for restoration of degraded biomes, mainly caused by human productive activities or due to the climate change.
ACKNOWLEDGEMENTS
The authors thank the staff of the Laboratorio de Cultivo de Tejidos Vegetales of the Universidad Ped-agógica y Tecnológica de Colombia and ECOPETROL, agreement 5211740 UPTC-ECOPETROL, for their support during the execution and development of this research. Also to Virginia Gutiérrez Pineda from the Departamento Administrativo de Ciencia, Tecnología e Innovación, COLCIENCIAS, and the Dirección de Investigaciones, DIN, of the Universidad Pedagógica y Tecnológica de Colombia. The research was financed by the program Joven Talento Colciencias-UPTC, agreement 701, 2013, Scholarship Jóvenes Investigadores e Innovadores 2013.
REFERENCES
1. Castaño C. Páramos y Ecosistemas alto andinos de Colombia en condición de hotspot y global climatic tensor. Bogotá: Instituto de Hidrología, Metereología y Estudios Ambientales, Ministerio de Medio Ambiente, IDEAM, PNUD; 2002.
2. Sturm H, Rangel O. Ecología de los páramos andinos. Una visión preliminar integrada. Bogotá: Instituto de Ciencias Naturales. Universidad Nacional de Colombia; 1985.
3. Van der Hammen T. Páramos. In: Informe nacional sobre el estado de la biodiversidad 1997-Colombia. Bogotá: Instituto de Investigación de Recursos biológicos Alexander von Humboldt; 1998.
4. Hofstede R, Coppus R, Mena P, Segarra P, Wolf J, Sevink J. The conservation status of tussock grass páramo in Ecuador. Ecotropicos. 2002;15(1):3-18.
5. Salinas C, Fuentes L, Hernández L. Caracterización de lepidópteros fitófagos asociados a la herviboría de frailejones en la microcuenca de la quebrada Calostros del Parque Nacional Natural Chingaza. Rev Mutis. 2013;3(1):1-22.
6. Díasgranados M. A nomenclator for the frailejones (Espeletiinae Cuatrec, Asteraceae). PhytoKeys. 2012;16:1-52.
7. Rivera D. Páramos de Colombia. Bogotá: Banco de Occidente; 2001.
8. Secretaria Distrital de Ambiente y Conservación Internacional Colombia. Formulación de la política de conservación de la biodiversidad en el distrito capital, su plan de acción y los lineamientos de conectividad ecológica. Diagnóstico sobre la conservación, conocimiento y uso de la biodiversidad del Distrito Capital de Bogotá, Colombia; 2010.
9. Díaz W, Torres C. Estudio básico de restauración vegetal en áreas de subpáramo degradadas de la Vereda Monquetiva-Guatavita. Colomb. For. 2001;7(14):116-23.
10. Ministerio de Agricultura y Desarrollo Rural. Colombia: Informe nacional para la conferencia técnica internacional de la FAO sobre los recursos fitogenéticos (Leipzig 1996). Bogotá: Ministerio de Agricultura y Desarrollo Rural; 1995.
11. Araque EJ, Bohórquez MD, Pacheco JC. Micropropagación de Espeletia paipana. Clonación de Frailejones. Tunja: Corporación Autónoma Regional de Boyacá, Corpoboyacá; 2014.
12. Bohórquez MD, Araque EJ, Pacheco JC. Propagación in vitro de Espeletia paipana S. Díaz y Pedraza, frailejón endémico en peligro de extinción. Actual Biol. 2016;38(104):23-36.
13. Murashige T, Skoog F. A revised medium for rapid growth and bio-assays with Tobacco tissue cultures. Physiol. Plant. 1962;15:473-97.
14. Folgueras M, Herrera L, Carrazana D. La contaminación microbiana en la micropropagación in vitro de las raíces y tubérculos tropicales. Libro de reportes cortos. Taller internacional BioVeg’. Ciego de Avila, Cuba; 2001.
15. Das M, Pal A. In vitro regeneration of Bambusa balcoa Roxb. Factors affecting changes of morphogenetic competence buds. Plant Cell Tissue Organ Cult. 2005;85(1):109-12.
16. Rodríguez M, Matehus J, Gersti A, Santana M. Identificación del agente causal de una bacteriosis en ñame (Dioscorea alata L.). Interciencia. 2008;33(7):1-11.
17. Murata Y, Katsura S, Obi I, Kakutani T. Alterations in Ca2+ binding on plasma membrane after adaptation to salt stress of tobacco cells in suspension. Plant Cell Physiol. 2000;41:1286-92.
18. Sánchez L, Rozo G, Montiel J. Cultivo in vitro de Gyrocarpus americanus (Hernandiaceae). Ibagué: Universidad del Tolima; 2012.
19. Sánchez M, Salaverría J. Control de la oxidación y la contaminación en el cultivo in vitro de fresa (Fragaria ananassa Duch.). UDO Agrícola. 2004;4:21-6.
20. Estrada C, Monasterio M. Comportamiento reproductivo de una roseta gigante, Espeletia spicata Sch. Bip. (Composite) del Páramo desértico. Ecotropicos. 1991;4(1):1-17.
21. Cavelier J, Machado JL, Valencia D, Montoya J, Laignelet A, Hurtado A, et al. Leaf demography and growth rates of Espeletia barclayana Cuatrec. (Compositae), a caulescent rosette in a Colombian paramo. Biotropica. 1992;24:52-63.
22. Berry P, Calvo R. Wind pollination, self-incompatibility and altitudinal shifts in pollination systems in the high Andean genus Espeletia (Asteraceae). Am J Bot. 1989;76:1602-14.
23. Sobrevila C. Effects of pollen donors on seed formation in Espeletia schultzii (Compositae) populations at different altitudes. Plant Syst Evol. 1989;166:45-67.
24. García Y, Freire M; Tejeda M, Reyes M. Germinación in vitro de semillas de Dendrocalamus strictus (Rosb.) Nees. Biotecnología Vegetal 2007;7(1):41-4.
25. Gielis J, Woods J, Oprins J. Micropropagation, synthetic seeds and germplasm storage of bamboos. 2004 [cited 2013 dic 26]. Available from: http://www.patentstorm.us/patents/6677154-description.html
26. Raggio M, Moro N, Busso C. Germinación in vitro de semillas de Encyclia adenocaula (La Llave & Lex.) Schltr (Orchidaceae). Phyton. 2008;77:203-15.
27. Guerra M, Pescador R, Dal Vesco L, Nodari R, Ducroquet J. In vitro morphogenesis in Feijoa sellowiana: somatic embryogenesis. Proc. Int. Sym. Myrtaceae. L. C. Donadio (Ed). Acta Hort. 1996;452:27-35.
28. Arditti J, Ghani A. Numerical and physical properties of orchid seeds and their biological implications. New Phytol. 2000;145(3):367-421.
29. Arditti J. Fundamentals of orchid biology. New York: Wiley-Interscience; 1992.
30. Arciga M. Germinación in vitro de Guarianthe aurantiaca Bateman (Orchidaceae) [dissertation]. Morelia, Mitch: Univ. Michoacana de San Nicolás; 2011.
31. Mayo A, Cázares J, De La Cruz E, Flores A. Germinación in vitro de semillas y desarrollo de plántulas de orquídeas silvestres de Tabasco. (Colección José N. Rovirosa, Biodiversidad, Desarrollo Sustentable y Trópico Húmedo). Tabasco México: Univ. Juárez Autónoma de Tabasco; 2010.
32. Brar J, Anand M, Sood A. In vitro seed germination of economically important edible bamboo Dendrocalamus membranaceus Munro. Indian J Exp Biol. 2013;51(1):88-96.
33. Vidales M. Regulación hormonal en las plantas. Función de relación en el reino Metafitas. Biología y Geología Argentina; 2003.
34. Gamborg OL, Wetter LR. Plant tissue culture methods. Ottawa: National research Council of Canada (NRC); 1975.
35. Roca W, Mroginski L. Cultivo de tejidos en la agricultura: Fundamentos y aplicaciones. Cali: Centro Internacional Agricultura Tropical, CIAT; 1991.
36. Hughes W. Ornamental species. Cloning agricultural plants via in vitro techniques. B.V. Conge (De). Boca Ratón, Florida: CRC Press; 1981.
37. Rache L, Pacheco J. Micropropagación de Espeletiopsis muiska (Cuatrecasas), frailejón del Parque Natural Ranchería-Boyacá, Colombia. Agron Colomb. 2009; 27(3):349-358.
38. Murashige T. Plant propagation through tissue cultures. Annu Rev Plant Physiol. 1974;25:135-66.
39. Ríos D, Avilés F, Sánchez M, Escobar R, Pereira G. Variación de la tasa de enraizamiento asociada al número de subcultivo y diámetro de microtallos de castaño Castanea sativa Mill. Agricultura Técnica (Chile). 2005;65(3):258-64.
40. Németh G. Induction of rooting. In: Baja Y. Biotechnology in agriculture and forestry. Berlin: Springer; 1986. p. 49-64.
41. Bennett IJ, McComb JA, Tonkin CM, McDavid DA. Alternating cytokinins in multiplication media stimulates in vitro shoot growth and rooting of Eucalyptus globulus Labill. Ann Bot. 1994;74:53-8.
42. Valencia M, Ramírez F. Notas sobre la morfología, anatomía y germinación de agraz (Vaccinium meridionale Sw). Agron Colomb. 1993;10(2):151-9.
43. Muñoz F. Propagación de flora endémica de páramo o en peligro de extinción en el Parque Natural del Cocuy. Boyacá: Parque Nacional del Cocuy; 2002.
44. Rangel JO, Sturm H. Consideraciones sobre la vegetación, la productividad y la artropofauna asociada a regiones paramunas de la cordillera oriental. In: Mora LE, Sturm H. Estudios ecológicos del páramo y del bosque altoandino cordillera oriental de Colombia. Tomo I. Bogotá: Academia Colombiana de Ciencias Exactas, Físicas y Naturales; 1995. p. 47-70.
Received in July 2015.
Accepted in March 2016.
Eyda J Araque-Barrera. Laboratorio de Cultivo de Tejidos Vegetales Bioplasma, Universidad Pedagógica y Tecnológica de Colombia, Colombia. E-mail: eyda.araque@uptc.edu.co


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}