










My SciELO
 Custom services
Custom servicesServices on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkBiotecnología Aplicada
On-line version ISSN 1027-2852
Biotecnol Apl vol.33 no.2 La Habana Apr.-June 2016
RESEARCH
P3, a monoclonal antibody capable to activate B-1a cells
P3, un anticuerpo monoclonal capaz de activar células B-1a
Darel Martínez, Lianet Cabrera, Ana María Hernández
Tumor Immunology Direction, Center of Molecular Immunology. 216 St. & 15th Ave, Havana, P.O.Box. 16040, Havana 11600, Cuba.
ABSTRACT
P3 is a murine IgM mAb that recognizes N-glycolylated gangliosides, glycolipids expressed on the cell membranes of most vertebrates. It also binds other self-antigens like sulfatides and has high homology with an antibody that recognizes myelin oligodendrocytes. Previously it was demonstrated that P3 mAb is able to activate an idiotypic cascade involving autologous anti-idiotypic B and T cells, in the absence of adjuvant or carrier protein, despite being a self-protein. The anti-P3 idiotype response is mediated not only by CD4+, but also by CD8+ T cells. B-1a cells are the dominant B cell population during the early phases of development, when the idiotypic connectivity is established. Germline natural antibodies secreted by these cells play fundamental roles fighting infections and keeping the homeostasis. In this work, we show that the P3 mAb was able to recognize B-1a cells from naïve BALB/c mice. This antibody induced the overexpression of the activation markers CD25, CD69 and CD86, and increased the secretion of IgM and a mixed pattern of IFN-γ, IL-4 and IL-10 cytokines by B-1a cells. The P3 mAb ability of activating the B-1a cells could contribute to its immunogenicity in the syngeneic model, further supporting its use as a tool to specifically target and stimulate B-1a cells.
Keywords: B-1a cells, idiotypic response, immunogenicity, monoclonal antibody.
RESUMEN
El P3 es un anticuerpo monoclonal murino de isotipo IgM, que reconoce a los gangliósidos N-glicolilados (un tipo de glicolípidos presente en la membrana celular de la mayoría de los vertebrados), a otros auto-antígenos como los sulfátidos y posee una alta homología con un anticuerpo que reconoce a la mielina de los oligodendrocitos. A pesar de ser una proteína autóloga, previamente se demostró que es capaz de activar una cascada idiotípica de células B y T CD4+ y CD8+ anti-idiotípicas en ausencia de adyuvantes o proteínas transportadoras. Las células B-1a son la población dominante durante las fases tempranas del desarrollo, cuando se establece la conectividad idiotípica, y los anticuerpos naturales de línea germinal secretados por estas células desempeñan un papel fundamental en la protección contra infecciones y el mantenimiento de la homeostasis. En este trabajo se demostró que el AcM P3 es capaz de reconocer a células B-1a de ratones BALB/c vírgenes. Este anticuerpo indujo en dichas células la expresión a altos niveles de los marcadores de activación CD25, CD69 y CD86, la secreción de IgM y un patrón mixto de las citocinas IFN-γ, IL-4 e IL-10. La capacidad del AcM P3 para activar células B-1a puede contribuir a su inmunogenicidad en el modelo singénico y, además, ser usada como una herramienta para direccionar específicamente contra las células B-1a y estimularlas.
Palabras clave: células B-1a, respuesta idiotípica, inmunogenicidad, anticuerpo monoclonal.
INTRODUCTION
P3 is a monoclonal antibody (mAb) of IgM isotype, which recognizes N-glycolylated gangliosides and sulfatides, both self-antigens in mice. It was also reported that the variable region of P3 is shared by the antibody A4ac, isolated from a mouse with experimental allergic encephalomyelitis, which recognizes myelin oligodendrocytes [1]. The P3 mAb triggers a strong anti-idiotypic response in the syngeneic BALB/c mice model, even in the absence of adjuvant
or carrier protein [2], which is not a common phenomenon [3]. Some authors have suggested that the IgM isotype or the presence of somatic hypermutations could be important factors to explain the immunogenicity of autologous immunoglobulins [4-6]. Curiously, the immunogenicity of the P3 mAb idiotype has been demonstrated in the absence of any constant domain [7] and the P3 mAb variable region is coded by germline genes [8]. Therefore, the intrinsic properties of the P3 mAb idiotype and its capacity to interact with immune cells could explain the capacity of P3 to induce a strong anti-idiotypic response.
Previously, we showed that both CD4+ and CD8+ T cells participate in the anti-P3 idiotypic response [9], in spite of the role of B cells remaining to be studied. Noteworthy, the variable region of the heavy chain (VH) of P3 mAb is germline and belongs to the Q52 (VH2) gene family, which was previously observed in autoantibodies against gangliosides and frequently used by CD5+ B-1 lymphocytes (B-1a) [8,10]. B-1a cells are the main B cell population of peritoneal and pleural cavities in mice [11], and represent the dominant B cell population during the early developmental phases, when the idiotypic connectivity is established [12].
B-1a cells are characterized by different levels of expression of typical B lymphocyte lineage surface molecules, as B220 (CD45R)low, IgMhigh, IgDlow, CD19 and CD43, but, unlike the B-2 cells, B-1a lack CD23 and express CD5. B-1a cells also differ from splenic B-2 cells in their antibody repertoire, developmental pathway, BCR signaling [13] and show a higher ca-pacity to activate T cells, preferentially to a Th1/Th17 phenotype [14-16]. A distinctive characteristic of B-1 cells is the spontaneous secretion of natural antibodies (nAbs). nAbs are usually of IgM isotype, coded by germline genes and have important functions both in the protection against pathogens and in the maintenance of homeostasis [17].
Therefore, this work was aimed to study the capacity of P3 mAb to recognize and activate B-1a cells. Our results showed that P3 mAb is able to recognize a high percentage of mouse peritoneal B-1a cells, stimulating this population in vitro to secrete IgM and a mixed cytokine pattern. The P3 mAb capacity to interact with B-1a cells may contribute to the high immunogenicity seen for this mAb and its capacity to activate CD4+ and CD8+ T cells in the syngeneic model.
MATERIALS AND METHODS
Animals
BALB/c mice, female of 6-8 weeks old, were purchased from the Center for Laboratory Animal Production (Cenpalab, Havana, Cuba). Animals were housed and bred in a barrier maintained room according to the guidelines stipulated by the Animal Subject Committee Reviews Board at the Center of Molecular Immunology (CIM, Havana, Cuba). The study was performed with the approval from CIM’s Institutional Animal Care and Use Committee and experiments were conducted in agreement with CIM’s institutional guidelines to avoid unnecessary animal suffering.
mAbs
The P3 mAb (IgM, k) recognizes NeuGc-containing gangliosides and sulphated glycolipids. It was isolated from BALB/c mice immunized with liposomes containing NeuGcGM3 [18]. The P3 mAb was purified from ascitic fluid by gel filtration chromatography using a Sephacryl S-300 high-resolution column (Pharmacia, Sweden). The column was equilibrated with PBS 1× (0.5 M NaCl, 2.7 mM KCl, 8 mM Na2HPO4, 1.5 mM KH2PO4; pH 7.4). The 6E4 (IgM, k) anti-mouse EGFR mAb, which is not immunogenic in the syngeneic model, was purified by the same methodology and used as isotype control in all the experiments. Both purified mAbs were pyrogen-free products according to results of the Gel-clot Endotoxin Testing (Pyrotell, USA).
ELISA for IgM quantification
Solid phase ELISA was performed using 96-well polystyrene MaxiSorp microtiter plates (Nunc, USA). Plates were coated with 10 μg/mL of goat anti-mouse IgM serum (Sigma, USA) in coating buffer (11 mM Na2CO3, 35 mM NaHCO3; pH 9.6) and incubated over night at 4 °C. Plates were blocked with PBS 1× (0.14 M NaCl, 2.7 mM KCl, 8 mM Na2HPO4, 1.5 mM KH2PO4, pH 7.4) plus 1 % BSA, for 1 h at 37 °C. Then, plates were incubated with serial dilutions of purified IgM mAbs from 1:300 to 1:24 300. Purified mouse polyclonal IgM antibodies (Sigma, USA) were used in a range of 0.8 to 100 ng/mL for quantification purposes. Alkaline phosphatase-conjugated goat anti-mouse IgM (Fcμ-specific; Jackson Immunoresearch Laboratories, USA) was used as secondary antibody at 0.03 μg/mL. The reaction was developed with 1 mg/mL p-nitro-phenyl phosphate substrate (Sigma, USA) in diethanolamine buffer (pH 9.8). Five washes in PBS 1× plus 0.05 % Tween 20 and 1 % BSA were carried out between each ELISA step. The absorbance was measured at 405 nm in an iMark ELISA microplate reader (Biorad, USA). The optical density (OD) values from the mAbs samples were analyzed using the Microsoft® Excel program, to determine a value of concentration by plotting the OD values into a commercial IgM standard curve (Sigma, USA).
Isolation of B-1a cells
Peritoneal washouts were obtained from naïve BALB/c mice and the B-1a cells were isolated by negative selection, using magnetic microbeads conjugated to a cocktail of biotin-conjugated antibodies against non-B-1a cells as first labeling reagent (B-1a Cell Biotin-Antibody Cocktail) and Anti-Biotin MicroBeads as secondary labeling reagent (Miltenyi Biotec, Germany), following the manufacturer’s instructions. The purity of the cell population was evaluated by flow cytometry, incubating the purified cells with anti-B220/FITC and anti-CD5/PECy5 (BD Bioscience, USA) for 20 min on ice. The binding was detected using a Gallios flow cytometer (8 Colors/2 Lasers configuration; Beckman Coulter, USA) and analyzed by the Kaluza 1.2 software (Beckman Coulter, USA). Binding was checked to be higher than 90 % in all cases.
Recognition of B-1a cells by P3 mAb
In order to study the capacity of P3 mAb to recognize peritoneal B-1a cells, peritoneal washouts were obtained from naïve BALB/c mice. Subsequently, 3 × 105 cells were incubated for 20 min on ice with anti- B220/FITC and anti-CD5/PECy5 (BD Bioscience), to identify B-1a cells, and 10 μg/mL of biotinylated P3 mAb or control IgM, followed by FITC-conjugated streptavidin (BD Biosciences, USA), all diluted in PBS 1× plus 1 % BSA. Binding was detected using a Gallios flow cytometer and analyzed by the Kaluza 1.2 software (both from Beckman Coulter, USA). In order to determine whether the immunization with P3 mAb increased the number of recognized cells, BALB/c mice received in the flank one subcutaneous dose of 50 μg of the antibody, in 200 μL of PBS 1×. Three days afterwards, mice were sacrificed, the peritoneal washouts obtained and peritoneal B-1a cells tested for recognition by P3 or the control mAb as described.
Activation of B-1a cells by P3 mAb
To determine whether P3 mAb was able to activate B-1a cells, 3 × 105 B-1a cells were isolated as previously described and cultured in RPMI-1640 medium (Gibco, USA), supplemented with 10 % fetal calf serum (FCS), sodium pyruvate, L-glutamine, 10 U/mL penicillin and 10 mg/mL streptomycin (Gibco, USA), for 72 h in the presence of 100 μg/mL of P3 mAb or an IgM isotype control (6E4 mAb, CIM, Cuba). The expression of B-1a cell activation markers was detected by flow cytometry, after the incubation of cells with anti-CD25/PE, anti-CD69 (M1.2F3)/PECy7 or anti-CD86/B7-2 (GL1)/PE for 20 min on ice, while the cell population phenotype was confirmed using anti-B220/FITC and anti-CD5/PECy5 (all con-jugates from BD Biosciences, USA).
Cytokine production was detected by intracellular staining. The cells were fixed and permeabilized, according to the manufacturer’s protocol, with BD Cytofix/Cytoperm Buffers and incubated with anti-IFN-γ/PE, anti-IL-4/PE and anti-IL-10/PE (BD Biosciences, USA). The staining was detected using a Gallios flow cytometer and analyzed by the Kaluza 1.2 software (Beckman Coulter, USA). In all cases, at least 10 000 events were acquired from lymphocytes in the B220+CD5+ region.
Detection of IgM secretion by ELISPOT
Peritoneal B-1a cells purified from BALB/c mice were incubated three days with 100 μg/mL of P3 mAb or control Ab, in the culture medium described before. After extensive washing 10 000 cells were transfer to ELISPOT plates (Multi-Screen-IP Plates, Millipore), coated with 10 μg/mL anti-mouse IgM Ab (SouthernBiotech). After six hours at 37 °C under CO2 atmosphere, the plates were washed with PBS 0.02 % Tween 20 and the pots of secreted antibodies were detected with 0.06 μg/mL alkaline phosphatase-conjugated goat anti-mouse IgM (Fcμ- specific; Jackson Immunoresearch Laboratories). The reaction was developed with 2 % dimethylformamide, 0.4 mM 5-bromo-4-chloro-3-indolyl phosphate and 0.36 mM p-nitro blue tetrazolium chloride substrate (KPL, USA), diluted in buffer 0.1 M NaHCO3/1.0 mM MgCl2, pH 9.8. The number of spots were determined using the Phoretix Expression software (NonLinear Dynamics, United Kingdom), filtering for circularity, peak height, volume and area.
Statistical analysis
In each experiment values were obtained in triplicate and figures shown one result representative of at least twice independent experiments. The differences between two groups were evaluated by Mann-Whitney U test. The differences were considered significant when p ≤ 0.05. All statistical tests were one-tailed, and conducted using SSPS for Windows version 19.0.0.1 software.
RESULTS
B-1a cells recognition by P3 mAb
The capacity of P3 mAb to recognize peritoneal B-1a cells obtained from naïve BALB/c mice was assessed by flow cytometry. P3 mAb specifically recognized up to 19 % of peritoneal B-1a cells, suggesting different clones are involved the interaction (Figure 1A and B). To determine whether a previous immunization would increase the percentage of recognized B-1a cells, the binding of P3 to peritoneal B-1a cells was analyzed three days after BALB/c mice were immunized with one dose of 50 μg of P3 mAb or the control IgM. There were no differences between the percentages of peritoneal B-1a cells recognized by P3 mAb from immunized or naïve mice (Figure 1 C and D). This suggested that P3 mAb immunization does not induce B-1a cells proliferation, which has been previously reported for other antigens recognized by this B cell population [19].
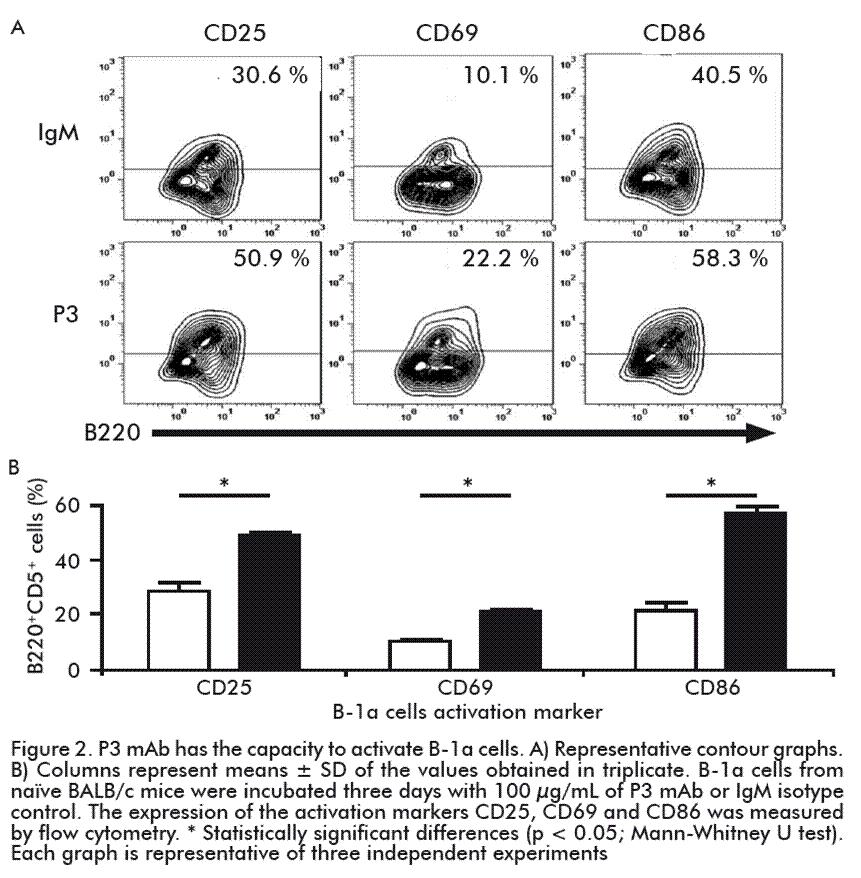
Evaluation of P3 mAb capacity to activate B-1a cells
Next, we wanted to know whether P3 mAb was able not only to recognize, but also activate B-1a cells.
With this aim, the expression of activation markers on the surface of naïve B-1a cells was tested after in vitro incubation with P3 mAb. As shown in figure 2, the culture of naïve peritoneal B-1a cells with P3 mAb during three days induced a higher percentage of B-1a cells expressing the activation markers CD25, CD69 and CD86 (1.6, 2.1 and 1.4 fold change, respectively), in comparison with the cells cultured with the control antibody. In addition, we tested IFN-γ, IL-10 and IL-4 cytokine production by naïve peritoneal B-1a cells by intracellular staining, after three days in culture with the P3 mAb. Compared to cells cultured with the control IgM, the presence of P3 mAb increased the percentages of B-1a cells producing all the cytokines tested, suggesting this mAb induces a mixed cytokine pattern (Figure 3).
Since a hallmark of B-1a cells is their capacity to spontaneously secrete IgM antibodies [17, 20], we evaluated whether the P3 mAb has the capacity to induce a higher level of IgM secretion by these cells. Peritoneal B-1a cells from naïve BALB/c mice were incubated three days with P3 mAb and the IgM secretion by B-1a cells was measured by ELISPOT. As is shown in figure 4, the P3 mAb was able to induce a significant increase in the number of spots of IgM secreting B-1a cells.
Discussion P3 is a monoclonal antibody with the ability to induce a strong anti-idiotypic response in the absence of adjuvant or carrier protein in the syngeneic model, which is not a frequently observed phenomenon [3, 21, 22]. It also recognizes other self-molecules like sulfatides [18]. Our group previously showed that P3 is able to activate idiotypic networks that involve not only B but also T cells [23]. In fact, it was also proved that CD4+ but also CD8+ T cells were indispensable for P3 to induce an anti-idiotypic response [9]. The variable region of P3 mAb is encoded by genes in the germline and belongs to the VHQ52 family (VH2) [8], a feature it shares with other anti-ganglioside antibodies [24-26]. This VH family is frequently used by B-1a cells, which recently were also proved to be able to secrete anti-NeuGcGM3 antibodies [27]. Since B-1a is the dominant B cell population in the neonatal period, when the idiotypic networks are established [17], and due to its ability to strongly activate T cells [14- 16], we studied whether P3 mAb was able to interact with B-1a.
P3 mAb was able to recognize in vitro up to 19 % of peritoneal B-1a cells, suggesting this antibody recognizes clones with different specificities. This could be explained by the existence of a regulatory idiotope in the variable region of P3 mAb, as was proposed by Pérez et al. [23]. The percentage of recognized B-1a cells did not increased with P3 mAb immunization, evidencing that this antibody does not induce B-1a cell proliferation in vivo. This was in agreement with previous reports for other antigens recognized by this B cell population [19].
Interestingly, in addition to recognizing peritoneal B-1a cells, P3 mAb was able to activate this population in vitro, without the mediation of any other cell population, increasing the percentage of cells expressing the activation markers CD25, CD69 and CD86.
Although the role of B cells in the immune response is usually framed in its capacity to secrete antibodies, B cells are professional antigen presenting cells [28-30]. Specifically B-1a cells constitutively express high levels of MHC-II and co-stimulatory molecules CD80 and CD86 [31], and several studies have demonstrated their ability to present antigens to T lymphocytes [15, 16]. The culture with P3 mAb also increased the percentage of B-1a cells producing IFN-γ, IL-4, and IL-10, a mixed cytokine pattern, reinforcing the idea that several B-1a clones are simultaneously activated. In our case, the percentage of IFN-γ+ B-1a cells was higher than IL-4+ or IL-10+ cells, suggesting the induction of a predominant Th1 response. This is consistent with previous reports showing a preferential polarization to Th1/Th17 by B-1a cells [14]. Our results suggest that B-1a cells could mediate the idiotypic interactions between B and T cells induced by this mAb in the syngeneic model.
The activation with P3 mAb in vitro also induced a higher percentage of B-1a cells secreting IgM, a fundamental role of these cells [32, 33]. Noteworthy, natural antibodies (nAbs) secreted by B-1a cells are the first barrier against pathogens, especially relevant before the establishment of a specific immune response [33]. Despite their low affinity, these antibodies can directly neutralize early replication of pathogens [34,35] or activate the complement cascade [36]. nAbs also keep the homeostasis by the removal of dead cells, oxidized and transformed epitopes [37,38]. Additionally, natural IgMs have been related with enhanced IgG response by B-2 cells [39,40]. nAbs frequently also cross-react with self-antigens, including other antibodies [41,42]. It would be important to study the specificities of the B-1a cells activated by P3 mAb. To our knowledge, this is the first report of a monoclonal antibody with the capacity to recognize, activate and stimulate B-1a cells to secrete nAbs. Further studies are required to confirm this potentiality in vivo. Moreover, the P3 mAb could be used as a tool to specifically stimulate this population in instances where B-1a cells and nAbs could be crucial, like immunodeficiency and atherosclerosis, among others [43-45].
CONFLICT OF INTERESTS STATEMENT
The authors declare the absence of conflict of interests.
REFERENCES
1. Libbey JE, Peterson LK, Tsunoda I, Fujinami RS. Monoclonal MOG-reactive autoantibody from progressive EAE has the characteristics of a natural antibody. J Neuroimmunol. 2006;173(1-2):135-45.
2. Vazquez AM, Perez A, Hernandez AM, Macias A, Alfonso M, Bombino G, et al. Syngeneic anti-idiotypic monoclonal antibodies to an anti-NeuGc-containing ganglioside monoclonal antibody. Hybridoma. 1998;17(6):527-34.
3. Ismaili J, Brait M, Leo O, Urbain J. Assessment of a functional role of auto-anti-idiotypes in idiotype dominance. Eur J Immunol. 1995;25(3):830-7.
4. Reitan SK, Hannestad K. The primary IgM antibody repertoire: a source of potent idiotype immunogens. Eur J Immunol. 2001;31(7):2143-53.
5. Reitan SK, Hannestad K. Immunoglobulin heavy chain constant regions regulate immunity and tolerance to idiotypes of antibody variable regions. Proc Natl Acad Sci U S A. 2002;99(11):7588-93.
6. Zhang X, Smith DS, Guth A, Wysocki LJ. A receptor presentation hypothesis for T cell help that recruits autoreactive B cells. J Immunol. 2001;166(3):1562-71.
7. Lopez-Requena A, Bestagno M, Mateo de Acosta C, Cesco-Gaspere M, Vazquez AM, Perez R, et al. Gangliosides, Ab1 and Ab2 antibodies III. The idiotype of anti-ganglioside mAb P3 is immunogenic in a T cell-dependent manner. Mol Immunol. 2007;44(11):2915-22.
8. Perez A, Lombardero J, Mateo C, Mustelier G, Alfonso M, Vazquez AM, et al. Immunogenetic analysis of variable regions encoding AB1 and gamma-type AB2 antibodies from the NeuGc-containing ganglioside family. Hybridoma. 2001;20(4):211-21.
9. Martinez D, Rodriguez N, Grinan T, Rondon T, Vazquez AM, Perez R, et al. P3 mAb: An Immunogenic Anti-NeuGcGM3 Antibody with Unusual Immunoregulatory Properties. Front Immunol. 2012;3:94.
10. Ben-Yehuda A, Szabo P, LeMaoult J, Manavalan JS, Weksler ME. Increased VH 11 and VH Q52 gene use by splenic B cells in old mice associated with oligoclonal expansions of CD5 + B cells. Mech Ageing Dev. 1998;103(2):111-21.
11. Margry B, Wieland WH, van Kooten PJ, van Eden W, Broere F. Peritoneal cavity B-1a cells promote peripheral CD4+ T-cell activation. Eur J Immunol. 2013;43(9):2317-26.
12. Elliott M, Kearney JF. Idiotypic regulation of development of the B-cell repertoire. Ann N Y Acad Sci. 1992;651:336-45.
13. Hayakawa K, Hardy RR. Development and function of B-1 cells. Curr Opin Immunol. 2000;12(3):346-53.
14. Zhong X, Gao W, Degauque N, Bai C, Lu Y, Kenny J, et al. Reciprocal generation of Th1/Th17 and T(reg) cells by B1 and B2 B cells. Eur J Immunol. 2007;37(9):2400-4.
15. Gao J, Ma X, Gu W, Fu M, An J, Xing Y, et al. Novel functions of murine B1 cells: active phagocytic and microbicidal abilities. Eur J Immunol. 2012;42(4):982-92.
16. Parra D, Rieger AM, Li J, Zhang YA, Randall LM, Hunter CA, et al. Pivotal advance: peritoneal cavity B-1 B cells have phagocytic and microbicidal capacities and present phagocytosed antigen to CD4+ T cells. J Leukoc Biol. 2012;91(4):525-36.
17. Baumgarth N. The double life of a B-1 cell: self-reactivity selects for protective effector functions. Nat Rev Immunol. 2011;11(1):34-46.
18. Vazquez AM, Alfonso M, Lanne B, Karlsson KA, Carr A, Barroso O, et al. Generation of a murine monoclonal antibody specific for N-glycolylneuraminic acid-containing gangliosides that also recognizes sulfated glycolipids. Hybridoma. 1995;14(6):551-6.
19. Morris DL, Rothstein TL. Abnormal transcription factor induction through the surface immunoglobulin M receptor of B-1 lymphocytes. J Exp Med. 1993;177(3):857-61.
20. Gronwall C, Silverman GJ. Natural IgM: beneficial autoantibodies for the control of inflammatory and autoimmune disease. J Clin Immunol. 2014;34 Suppl 1:S12-21.
21. Bogen B, Jorgensen T, Hannestad K. Recognition of lambda 1 and lambda 2 murine light chains by carrier-specific isologous T helper cells; effect of L-H chain assembly. Eur J Immunol. 1983;13(5):353-9.
22. Hernandez AM, Rodriguez M, Lopez- Requena A, Beausoleil I, Perez R, Vazquez AM. Generation of anti-Neu-glycolyl-ganglioside antibodies by immunization with an anti-idiotype monoclonal antibody: A self versus non-self-matter. Immunobiology. 2005;210(1):11-21.
23. Perez A, Mier ES, Vispo NS, Vazquez AM, Perez Rodriguez R. A monoclonal antibody against NeuGc-containing gangliosides contains a regulatory idiotope involved in the interaction with B and T cells. Mol Immunol. 2002;39(1-2):103-12.
24. Zenita K, Hirashima K, Shigeta K, Hiraiwa N, Takada A, Hashimoto K, et al. Northern hybridization analysis of VH gene expression in murine monoclonal antibodies directed to cancer-associated ganglioside antigens having various sialic acid linkages. J Immunol. 1990;144(11):4442-51.
25. Weng NP, Ritter E, Yucel E, Zhang D, Ritter G, Marcus DM. Specificity and structure of murine monoclonal antibodies against GM1 ganglioside. J Neuroimmunol. 1994;55(1):61-8.
26. Lopez-Requena A, Mateo De Acosta C, Vazquez AM, Perez R. Immunogenicity of autologous immunoglobulins: principles and practices. Mol Immunol. 2007;44(11):3076-82.
27. Rodriguez-Zhurbenko N, Rabade-Chediak M, Martinez D, Grinan T, Hernandez AM. Anti-NeuGcGM3 reactivity: a possible role of natural antibodies and B-1 cells in tumor immunosurveillance. Ann N Y Acad Sci. 2015;1362:224-38.
28. van Rooijen N. Direct intrafollicular differentiation of memory B cells into plasma cells. Immunol Today. 1990;11(5):154-7.
29. Kelsoe G. Life and death in germinal centers (redux). Immunity. 1996;4(2):107-11.
30. Gulbranson-Judge A, Casamayor-Palleja M, MacLennan IC. Mutually dependent T and B cell responses in germinal centers. Ann N Y Acad Sci. 1997;815:199-210.
31. Vigna AF, Godoy LC, Rogerio de Almeida S, Mariano M, Lopes JD. Characterization of B-1b cells as antigen presenting cells in the immune response to gp43 from Paracoccidioides brasiliensis in vitro. Immunol Lett. 2002;83(1):61-6.
32. Ehrenstein MR, O’Keefe TL, Davies SL, Neuberger MS. Targeted gene disruption reveals a role for natural secretory IgM in the maturation of the primary immune response. Proc Natl Acad Sci U S A. 1998;95(17):10089-93.
33. Baumgarth N, Tung JW, Herzenberg LA. Inherent specificities in natural antibodies: a key to immune defense against pathogen invasion. Springer Semin Immunopathol. 2005;26(4):347-62.
34. Ochsenbein AF, Fehr T, Lutz C, Suter M, Brombacher F, Hengartner H, et al. Control of early viral and bacterial distribution and disease by natural antibodies. Science. 1999;286(5447):2156-9.
35. Baumgarth N, Chen J, Herman OC, Jager GC, Herzenberg LA. The role of B-1 and B-2 cells in immune protection from influenza virus infection. Curr Top Microbiol Immunol. 2000;252:163-9.
36. Jayasekera JP, Moseman EA, Carroll MC. Natural antibody and complement mediate neutralization of influenza virus in the absence of prior immunity. J Virol. 2007;81(7):3487-94.
37. Chou MY, Fogelstrand L, Hartvigsen K, Hansen LF, Woelkers D, Shaw PX, et al. Oxidation-specific epitopes are dominant targets of innate natural antibodies in mice and humans. J Clin Invest. 2009;119(5):1335-49.
38. Kaveri SV, Silverman GJ, Bayry J. Natural IgM in immune equilibrium and harnessing their therapeutic potential. J Immunol. 2012;188(3):939-45.
39. Boes M, Esau C, Fischer MB, Schmidt T, Carroll M, Chen J. Enhanced B-1 cell development, but impaired IgG antibody responses in mice deficient in secreted IgM. J Immunol. 1998;160(10):4776-87.
40. Baumgarth N, Herman OC, Jager GC, Brown LE, Herzenberg LA, Chen J. B-1 and B-2 cell-derived immunoglobulin M antibodies are nonredundant components of the protective response to influenza virus infection. J Exp Med. 2000;192(2):271-80.
41. Jeong HD, Teale JM. Contribution of the CD5+ B cell to D-proximal VH family expression early in ontogeny. J Immunol. 1990;145(8):2725-9.
42. Casali P, Kasaian MT, Haughton G. B-1 (CD5 B) cells. In: Coutinho A, Kazatchkine MD, editores. Autoimmunity Physiology and Disease New York: Wiley-Liss Inc.; 1994. p. 57.
43. Bayry J, Lacroix-Desmazes S, Donkova- Petrini V, Carbonneil C, Misra N, Lepelletier Y, et al. Natural antibodies sustain differentiation and maturation of human dendritic cells. Proc Natl Acad Sci U S A. 2004;101(39) :14210-5.
44. Hamanova M, Chmelikova M, Nentwich I, Thon V, Lokaj J. Anti-Gal IgM, IgA and IgG natural antibodies in childhood. Immunol Lett. 2015;164(1):40-3.
45. Rothstein TL. Natural Antibodies as Rheostats for Susceptibility to Chronic Diseases in the Aged. Front Immunol. 2016;7:127.
Received in June, 2016.
Accepted in June, 2016.
Ana María Hernández. Tumor Immunology Direction, Center of Molecular Immunology. 216 St. & 15th Ave, Havana, P.O.Box. 16040, Havana 11600, Cuba. E-mail: anita@cim.sld.cu.


{kind=link}
{kind=link}
{kind=link}
{kind=link}