










My SciELO
 Custom services
Custom servicesServices on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkBiotecnología Aplicada
On-line version ISSN 1027-2852
Biotecnol Apl vol.33 no.3 La Habana July.Sept. 2016
FOCUS
Methodological procedure to obtain rice (Oryza sativa L.) cultivars resistant to Pyricularia grisea Sacc. with good agronomic performance
Procedimiento metodológico para la obtención de cultivares de arroz (Oryza sativa L.) resistentes a Pyricularia grisea Sacc. y con buen comportamiento agronómico
Noraida de Jesús Pérez León
Unidad Científico Tecnológica de Base (UCTB) Los Palacios, Instituto Nacional de Ciencias Agrícolas, INCA. Carretera a Tapaste, Km. 3 1/2 San José de las Lajas, CP 32700, Mayabeque, Cuba.
ABSTRACT
Blast is caused by the fungus Pyricularia grisea, and it is the main pathological problem affecting rice cultivation yields in Cuba. Therefore, a methodological procedure was established to select rice cultivars resistant to P. grisea and showing good agronomic performance, through the use of in vitro anther culture of F2 plants. Blast-resistant progenitors were crossed up and anthers of F2 plants were cultured in vitro for callus formation and subsequent plant regeneration. The first and second generations of plants were evaluated for agronomic traits, and the second generation for resistance to Blast. Selected lines were further challenged against aggressive haplotypes isolated in Cuba under controlled conditions of natural infection and high pressure of the pathogen at the ‘Caribe’ hotspot, and the resistant ones were characterized attending to 51 morphoagronomic descriptors. Four resistant cultivars and other six of good agronomic performance were selected as parents for the designed breeding program. Plant genotype and NL culture medium were crucial for in vitro anther culture success, with greater callus formation, and three combinations showed the best responses. In the second generation, nine lines were identified combining resistance and high yields by natural infection under field conditions. Of them, seven were resistant to inoculation with pathogen haplotypes isolated in Cuba. Moreover, three were resistant under field conditions against the P. grisea variability at the ‘Caribe’ hotspot. The methodological procedure reported shortens the cultivar improvement cycle, creating variability faster and with adequate fixation of relevant characters. One obtained line, the ‘Anays LP-14’, was registered in the Commercial Register of Cuban varieties for commercial exploitation.
Keywords: rice improvement, Pyricularia grisea, Blast, in vitro anther culture, breeding program.
RESUMEN
La enfermedad conocida como el tizón del arroz (Blast) es causada por el hongo Pyricularia grisea, que provoca la principal afectación de ese cultivo en Cuba. Se estableció una metodología para la selección de cultivares de arroz, resistentes a P. grisea y de buen comportamiento agronómico, obtenidos mediante el cruce de progenitores resistentes a Blast y el cultivo in vitro de anteras de plantas F2, con la posterior formación de callos y su regeneración. La primera y segunda generaciones de plantas fueron evaluadas según 51 descriptores morfoagronómicos, además del comportamiento frente a Blast en la segunda. Se seleccionaron cuatro cultivares resistentes a P. grisea y otros seis de buen comportamiento agronómico como progenitores, para el programa de mejora. El genotipo y el medio de cultivo NL fueron determinantes para el éxito del cultivo in vitro de las anteras, con una mayor formación de callos, y tres combinaciones mostraron una respuesta superior. En la segunda generación se identificaron nueve líneas bajo infección natural de P. grisea en campo, siete de ellas resistentes a aislamientos de haplotipos cubanos del patógeno y tres mostraron resistencia en campo frente a toda la variabilidad de P. grisea existente en la localidad ‘Caribe’, con alta presión del patógeno. El procedimiento metodológico acortó el ciclo de mejora, creó variabilidad y logró una rápida fijación de los caracteres de las líneas resistentes y altamente productivas. Una de esas líneas, la ‘Anays LP-14’ fue inscrita en el Registro Comercial de Variedades Cubanas.
Palabras clave: mejora del arroz, Pyricularia grisea, tizón del arroz, cultivo in vitro de anteras, cruzamirento genético.
INTRODUCTION
Rice (Oryza sativa L.) is one of the most economically relevant grain crops worldwide. Along with wheat, meat and fish, rice forms part of the basic human diet, with 75 % of the world population having it in their daily diet and, in some cases, overcoming the consumption of other cereals [1, 2].
In Cuba, it is the main source of carbohydrates in the diet, with an estimated consumption of 670 000 tons per year. So far, domestic production meets only about 50 % of the needs, forcing to import the rest. The average agricultural yield remains close to 3 t·ha-1, below the world average, which is motivated by different causes, among them: planting outside the optimal season, deficient cultivation care, lack of irrigation, continuous soil salinization and pest damages [3].
Among the most significant rice crop pests, we found the fungus Magnaporthe grisea Barr (Pyricularia grisea Sacc), causing Blast, a disease considered the most devastating worldwide and of a widespread distribution [4]. It affects leaves, stems and knots of the plant and the different parts of the panicles and grains, ultimately leading to a significant reduction of agricultural yields [5] and sometimes to the complete loss of the harvest. Therefore, this disease represents a serious limitation for rice production as the main rice crop pest in Cuba [6].
Notably, Cuban rice production was sustained for a long time by a single cultivar, J-104, which was shown to be susceptible to Blast. This led to severely reduced yields and made necessary to apply fungicide treatments to reduce the incidence of the fungus, further raising the costs for crop production and contributing to environmental pollution. Even the current strategy of planting from two-to-four cultivars by rice-growing area does not guarantee the absence of infection, since cultivars are not resistant to the disease. Therefore, the problem persists and the development of resistant cultivars is still a relevant economic route for disease control, of lower environmental impact [7].
On the other hand, it is known that several genes are involved in the inheritance of resistance to this disease and against the high variability displayed by the pathogen, this last promoting the appearance of new haplotypes able to overcome the resistance of cultivars [8]. This implies that, the adequate site with high pressure and diversity of the pathogen diversity must be identified, for the selection of plant parental and resistant lines, associated with favorable environmental conditions for plant development, supporting the selection of rice lines with a large array of genes as to enable resistance against the pathogen both steadily and inheritably [7]. Moreover, in any breeding program for pest resistance, a controlled genetic resistance source is required, to establish suitable conditions for pathogen infection, in order to guarantee an effective selection and to choose the appropriate breeding method.
Among them, the in vitro anther culture of F2 plants offers many advantages, since the population obtained from this combination of improvement methods (hybridizations and in vitro culture) is representative of the genetic variability of the F2 population, and the plants obtained in vitro are genetically homozygotic. This reduces the time for obtaining new cultivars, saves financial and material resources, increases the efficiency of selection and facilitates the identification of superior individuals [9-11].
In Cuba, breeding programs for waterlogging agro-ecosystems have been based mainly on hybridizations, but unfortunately, the obtained cultivars were susceptible to Blast, in spite of showing potentiality for high production yields [12-14]. Moreover, the ‘Caribe’ site, belonging to the Agroindustrial Rice Complex ‘Los Palacios’ in the Pinar del Río province, has historically demonstrated a high incidence of the disease [3] and, therefore, it has been selected for crop breeding to develop cultivars resistant to Blast. Given the issues raised, a methodological procedure was developed, aimed to obtain rice cultivars resistant to P. grisea infection and showing good agronomic performance through in vitro anther culture of F2 plants, followed by successive selection steps of the lines obtained.
METHODOLOGICAL PROCEDURE
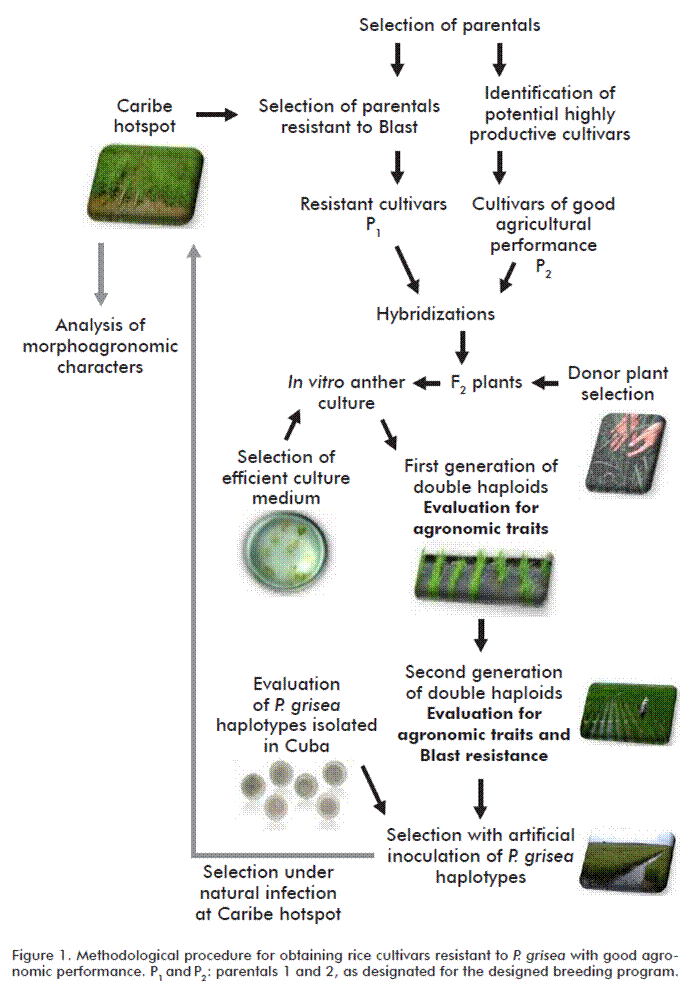
The methodological procedure developed (Figure) starts with the selection of Blast resistant parentals [15]. They were established by evaluating commercial and pre-commercial cultivars for performance when subjected to the natural infection with the disease in the ‘Caribe’ hotspot and using the methodology validated in Cuba [16]. Cultivars showing high production potentiality in different cultivation areas dedicated to rice production were identified [17].
Subsequently, the resistant parentals of good agricultural performance were subjected to a breeding program, by hybridization according to a statistical genetic design, and the F1 seeds obtained from the crossings were planted for reproduction.
In vitro anther culture of F2 plants was applied as a second step, selecting for donor plants which anthers were cultured, also including the optimization of the culture medium for callus formation and the regeneration of green plants [18].
Different factors influence the double-haploid production, but the genotype is regarded as the most influential one in callus formation, determining the ability of calluses to differentiate into plants at the adequate green:albino plant ratio and the ploidy of the regenerated explants. Moreover, the number of isogenic lines to be produced is also influenced by the genetic background of the cultivar itself, since a higher number of isogenic lines can be obtained from many japonica rice cultivars as compared to the low production of calluses and the low regeneration efficiency of green plants from most indica cultivars [19]. In this regard, there is evidence that the response of cultivars to callus induction and plant regeneration is inherited and controlled by nuclear genes, which can be transferred to recalcitrant lines through genetic recombination [20].
Although initiation of microspores division may be independent of nutritional adjuncts, these are required for subsequent divisions, which lead to callus formation and differentiation of these cells in embryos and plants. Several laboratories have tried to optimize the anther culture rice medium composition, but their findings differ from each other due to the influence of genotype, the state of pollen development and the temperature pretreatment of anthers. Significantly, medium composition is still relevant, since its changes produce variations in the frequency of induction for the same genotype [21].
Then, several selection cycles were implemented, starting with the first generation of plants obtained in vitro, which were evaluated for agronomic traits. In the second generation, a simultaneous selection of two characters was made with independent criteria: cultivars resistant to Blast, in leaf and panicle neck, by natural field infection and productive potential [22].
The lines combining resistance to Blast and good agronomic performance were evaluated under controlled conditions against P. grisea aggressive haplotypes isolated in Cuba [23], and under natural infection at the ‘Caribe’ site, under high pathogen pressure. Finally, the lines displaying resistance were further characterized for morphoagronomic characters [24].
Blast is considered a complex disease due to the pathogenic variability and the speed with which the fungus overcomes the resistance of the rice plant [25]. With the application of molecular techniques, the high diversity of population pathotypes was grouped in a smaller group of lineages according to their genetic similarity [26].
In the Philippines, in studies conducted with pathogen populations, 71 haplotypes from six genetic lineages were identified and a consistent incompatibility was evidenced for most host-lineage combinations. This led to the assumption that, although some resistance genes can be easily overcome by a given pathogen lineage, these same resistance genes can be a good barrier to other lineages of the population, and viceversa [8, 27].
In Cuba, the first reports on the identification of circulating P. grisea fungus isolates date back to 1984 [28]. Later on, works on the genetic diversity of the fungus evaluated at two Cuban locations (‘Los Palacios’ and ‘Sur del Jíbaro’) showed that the pathogen population consisted of four closely-related lineages with a high number of haplotypes (up to 134). The first location showed the highest genetic diversity and the largest number of genetic lineages, in agreement with the highest incidence reported for this region in Cuba [29].
Additionally, the great pathogenic variability of P. grisea is one of the major constraints in breeding. Thus, the characterization of lineages present in each country contributes significantly to the establishment of breeding strategies, based on the theory of exclusion lineages. Such strategies consider the coexistence of different lineages groups in each rice growing region, with progenitors needing to be selected to provide resistance genes to such groups [7].
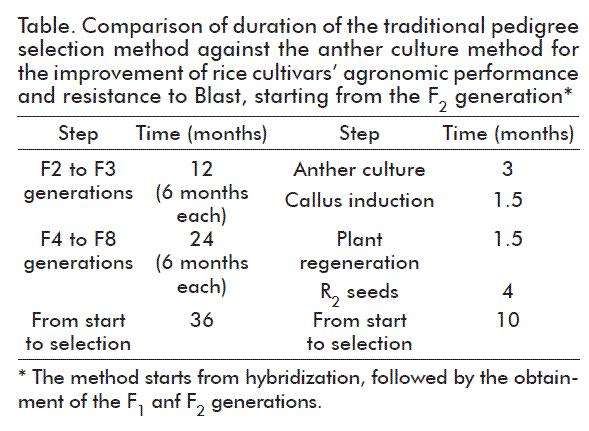
After the implementation of the methodology described here, homozygous Blast-resistant lines were identified after sowing of the F2 seed (Table), with good agronomic performance in just 10 months. Therefore, the efficiency of the breeding method was confirmed. This result was equivalent to that attained with the selection method by pedigree, traditionally used in rice cultivation in Cuba, further accounting for the same stability usually achieved only after seven-to-eight generations of self-pollination with one or two annual crops during three-to-six working years [30].
A population of double haploids from anther culture of F2 plants accounted for the genetic variability of the entire F2 population, with the advantage that plants are genetically homozygotic. Consequently, the time to obtain the cultivars is reduced, providing considerable financial and material savings, not requiring large planting areas or any other production costs. It also increases the efficiency for selection, both qualitatively and quantitatively and facilitates the selection of superior cultivars [9-11, 31-33].
The introduction of in vitro anther culture has given varied results in different countries. For instance, in Venezuela [34], cultivars adapted to their cultivation conditions were obtained through anther culture, with immature embryos reported. In Chile, the F1 and F2 generations from triple crossings showed high sterility, whereas 941 R2 lines were obtained by anther culture, and further evaluated and selected for cold tolerance, better quality and early maturity. By these procedures, new lines of higher yields were obtained in considerably shorter periods [35]. In Colombia [36], fertile homozygotic lines were produced from interspecific Oryza sativa L. × Oryza barthii Chev and Oryza glaberrima × Oryza rufipogon Griff Steudt crossings, as new sources for pest resistance.
Additionally, durable Blast resistance requires: i) the characterization of: the genetic structure of P. grisea populations; ii) the diversity and frequency of avirulence/virulence pathogen genes and knowledge on its influence on resistance genes; iii) the identification and incorporation of combinations of resistance genes in rice germplasm and, once incorporated, the evaluation of the populations by controlled inoculations under field conditions, followed up through the use of molecular markers [37].
As previously mentioned, the evaluation of cultivars under field conditions requires identifying the hotspot site, characterized by a high pressure of the disease and high pathogen diversity which is associated with favorable environmental conditions for plant development. In that area, rice lines can be selected that combine a large set of resistance genes, further increasing their chances for inheritance and persistence. In Cuba, the ‘Caribe’ site, belonging to the Rice Agricultural Complex Los Palacios, is recommended for rice crop genetic breeding procedures to develop cultivars with Blast resistance, given its highest genetic diversity for two pathogen populations [29]. This region has been historically reported of a high incidence of the disease [3].
In order to improve the methods to obtain Blast resistant cultivars, other groups compared the reactions to P. grisea on the leaf and neck panicle of plant populations obtained by hybridizations and anther culture. They concluded that the latter method increases the efficiency for selection, due to a higher additive variation, the absence of dominance, an absence of intrafamiliar variation and competition between plants. All these facilitate the identification of superior cultivars attending to the selection in early generations of a pedigree program, and heritability values suggested that genetic breakthrough could be achieved in just a few generations [38].
Regarding resistance genes, P. grisea resistant lines were selected from the crossing progeny of two isogenic lines carrying the resistance genes Pi-1 and Pi-2, those lines shown to be incompatible with the fungus genetic lineages. This suggested the possibility to obtain lines which could exclude the entire pathogen population and displaying sustained resistance [39].
In assessing the progeny of a crossing between a cultivar carrying the Pi-ta gene with one that did, that gene was not found in susceptible individuals, suggesting that it acts as a dominant gene [40]. In later works, the Pi-ta gene was found in a group of American cultivars, as well as in the cultivar ‘Tetep’ of Vietnamese origin [41] which also carried the Pi-1 and Pi-kh genes [42]. Meanwhile, while growing African native ‘Moroberekan’ japonica type, the resistance found was attributed to the presence of a group of major resistance genes [43, 44]. Resistant lines were obtained from crossings with cultivar ‘IR 759-54-2-2’ [45], but neither the genes responsible for resistance nor the underlying mechanism acting could be identified. In fact, CIAT studies have shown that the combination of resistance genes Pi-1 (chromosome 11), Pi-2 (chromosome 6), and Pi-33 gene (chromosome 8) confer resistance to many populations of Latin America P. grisea [8, 46].
Notably, the methodological procedure developed [18] rendered three recombinant lines, regenerated from F2 plants anther culture, which apparently inherited the genes present in the resistant parental used (‘IR 759-54-2-2’ ‘Tetep’, ‘2077’ and ‘Moroberekan’). They confered Blast resistance, as evaluated in leaf in the vegetative phase and neck panicle at the flowering stage, supported for several years under a high pressure disease and pathogen diversity under field conditions.
CONCLUSIONS
Consistently, rice parentals for a rice breeding program were identified. The genotype and the culture medium used were crucial to the success of F2 plants in vitro anther culture, with the NL medium aiding to induce an increased callus formation.
Nine lines were identified, which were simultaneously resistant to the pathogen and produced high yields in the second generation obtained lines and after their evaluation under natural infection P. grisea in field conditions. Seven of those lines showed resistance when inoculated with isolates of Cuban haplotypes of the pathogen. Moreover, three three of the lines showed resistance under field conditions against all the existing variability of P. grisea in the ‘Caribe’ location, a site established as ideal for Blast evaluation in Cuba.
In summary, a methodological procedure was obtained and tested to shorten the breeding cycle in relation to crossbreeding, which creates variability and manages a quick fixation of the characters through the in vitro anther culture of F2 plants. This procedure also supports the adequate selection of lines for resistance to P. grisea and main agronomic characters, one of such lines, the ‘Anays LP-14’ which was obtained by this procedure, been already registered in the Commercial Register of Cuban varieties for crop improvement.
REFERENCES
1. Alvarez RM, Pérez M, Reyes E, Moreno OJ, Delgado N, Torrealba GT, et al. Evaluación comparativa de híbridos y variedades de arroz en los llanos centroccidentales de Venezuela. Agronomía Trop. 2008;58(2):101-10.
2. Méndez P. Arroz: ¿estabilidad o nueva alza de los precios mundiales? Info@rroz. 2011 Feb;(84):1.
3. MINAG. Instructivos Técnicos para el cultivo del arroz. La Habana: Instituto de Investigaciones del Arroz; 2013.
4. Castejón-Muñoz M. The effect of temperature and relative humidity on the airbone concentration of Pyricularia oryzae spores and the development of rice blast in southern Spain. Spanish J Agric Res. 2008;6(1):61-9.
5. Rodríguez AT, Ramírez MA, Cárdenas RM, Rivero D, Cruz A, Bautista S. Actividad antifúngica in vitro e in vivo del extracto acuoso de hojas de Parthenium hysterophorus L. sobre Pyricularia grisea Sacc. Revista Científica UDO Agrícola. 2012;12(4):839-44.
6. Cárdenas RM, Mesa S, Polón R, Pérez N, Cristo E, Fabré L, et al. Relación entre la incidencia de la piriculariosis (Pyricularia grisea Sacc.) del arroz (Oryza sativa Lin.) y diferentes variables climáticas en el Complejo Agroindustrial Arrocero Los Palacios. Cult Trop. 2010;31(1):14-8.
7. Zambrano AY, Vegas A, Cardona R, Gutiérrez Z, Demey JR. Estructura genética y diversidad de linajes de Pyricularia grisea en la zona arrocera venezolana. Interciencia. 2006;31(1):62-6.
8. Livore A. Desarrollo de una estrategia para la obtención de resistencia durable a Pyricularia grisea en Arroz en el Cono Sur. Informe técnico final – Proyecto FONTAGRO - convenio iica/bid ftg/rf-99-02-rg, INTA. San Isidro: Instituto Interamericano de Cooperación para la Agricultura (IICA); 2005 [cited 2016 Jun 15]. Available from: http://www.procisur.org.uy/online/DOCS/pyricularia22.pdf.
9. Ramakrishnan SH, Saravanan S, Anandakumar CR, Kannanbapu JR. In vitro androgénesis in rice (Oryza sativa L.). Asian J Plant Sci. 2005;4(6):600-2.
10. Chen C, Xiao H, Zhang W, Wang A, Xia Z, Li X, et al. Adapting rice anther culture to gene transformation and RNA interference. Sci China C Life Sci. 2006;49(5):414-28.
11. MINAG-Perú. Arroz. Producción Mundial. Portal Agrario. Cultivos de importancia nacional. 2016 [cited 2016 Jan 10]. Available from: http://www.minag.gob.pe/arroz/produccion.html.
12. Suárez E, Pérez R, Alfonso R, Mesa H, Deus JE, Cruz F, et al. Nuevas variedades de ciclo corto en Cuba. Arroz en las Américas. 1997;18(1):2.
13. Pérez N, Castro RI. A new short cycle rice variety: INCA LP-5. Cult Trop. 2000;21(4):55.
14. IIArroz. Política varietal para el cultivo del arroz. Anexos al Instructivo Técnico del Arroz. La Habana: Instituto de Investigaciones del Arroz (IIA); 2001.
15. Pérez N, González MC, Cárdenas RM, Castro RI. Selección de progenitores resistentes a la Piriculariosis para los programas de cruzamientos en arroz. Cult Trop. 2009;30(3):47-9.
16. Cárdenas RM, Cordero V, Pérez N, Cristo E, Gell I. Utilización de una nueva metodología para la evaluación de arroz ante la infección producida por el hongo Pyricularia grisea. Cultivos Tropicales 2000;21(1):63-6.
17. Pérez N, González MC, Castro RI, Aguilar M. Evaluación de cultivares de arroz en diferentes zonas de producción arrocera de “Los Palacios”, Pinar del Río, para su utilización en programas de mejoramiento. Cult Trop. 2016;37(1):116-23.
18. Pérez N, González MC, Castro RI, Aguilar M. Nuevos genotipos de arroz resistentes a la Piriculariosis obtenidos por cultivo de anteras. Rev Colomb Biotecnol. 2012;XIV(1): 256-70.
19. Silva TD, Ratnayake WJ. Anther culture potential of indica rice varieties, kurulu thuda and Bg 250. Trop Agric Res Ext. 2009;12(2):53-6.
20. Bagheri N, Jelodar NB. Combining ability and heritability of callus induction and green plant regeneration in rice anther culture. Biotechnology. 2008;7(2):287-92.
21. Herath HM, Bandara DC, Samarajeewa PK. Effect of culture media for anther culture of indica rice varieties and hybrids of indica and japonica. Trop Agric Res Ext. 2007;10:17-22.
22. Pérez NJ, González MC, Alvarez M, Aguilar M, Castro RI. Estudios preliminares referidos a la herencia de la resistencia a la Piriculariosis de líneas isogénicas de arroz obtenidas por cultivo de anteras. Cultivos Tropicales 2013;34(1):60-5.
23. Pérez N, González MC, Marquez R, Castro RI, Aguilar M. Utilización de haplotipos de Pyricularia grisea Sacc. aislados en Cuba para la selección de cultivares de arroz resistentes a la Piriculariosis. Cult Trop. 2015;36(1):129-33.
24. Pérez N, González MC, Castro RI, Aguilar M. Caracterización agronómica de tres cultivares de arroz (Oryza sativa L.) obtenidos mediante el cultivo in vitro de anteras. Cult Trop. 2016;37(1):110-5.
25. Anjos LM, Santos GR, Dias-Neto JJ, Oliveira WF, Castro Neto MD. Identification of physiological races of Magnaporthe grisea in areas of rice irrigated in the State of Tocantins. Trop Plant Pathol. 2009;34(3):80.
26. Levy M, Romao J, Marchetti MA, Hamer JE. DNA Fingerprinting with a Dispersed Repeated Sequence Resolves Pathotype Diversity in the Rice Blast Fungus. Plant Cell. 1991;3(1):95-102.
27. Zeigler RS, Cuoc LX, Scott RP, Bernardo MA, Chen DH, Valent B, et al. The relationship between lineage and virulence in Pyricularia grisea in the Philippines. Phytopathology. 1995;85:443-51.
28. Fabregat M. Aspectos bioecológicos y control de Pyricularia oryzae Cav. en el arroz. La Habana: Instituto Superior de Ciencias Agropecuarias de la Habana, Facultad de Agronomia; 1984.
29. Fuentes JL. Estructura y diversidad genética de poblaciones cubanas del hongo Pyricularia grísea [Tesis de Maestría]. La Habana: Centro de Aplicación Tecnológica y Desarrollo Nuclear (Ceaden); 1998.
30. Pérez N. Obtención de cultivares de arroz (Oryza sativa L.) resistentes a Pyricularia grisea Sacc. con buen comportamiento agronómico. [Tesis de Doctorado]. San José de las Lajas: Instituto Nacional de Ciencias Agrícolas; 2012.
31. Jiang J, Gibbons JW, Moldenhauer KAK, Johnson VA, Lee FN. The use of anther culture for rice variety development in Arkansas. AAES Research Series. 2004;(517):49-57.
32. Pérez-Almeida I. Aplicaciones Biotecnológicas en el Mejoramiento del Arroz. Revista Digital CENIAP HOY. 2004[cited 2014 Oct 31];(6). Disponible en: http://www.ceniap.gov.ve/ceniaphoy/articulos/n6/arti/perez_almeida_i/art/perez_almeida_i.htm.
33. Purwoko BS, Dewi IS, Khumaida N. Rice anther culture to obtain doubled-haploids with multiple tolerances. Asia Pac J Mol Biol Biotechnol. 2010;18(1):55-7.
34. Casale I. Laboratorio de Mejoramiento de Plantas. Caracas: Instituto de Zoología Tropical. Facultad de Ciencias. 2006 [cited 2006 Jan 10]. Available from: http://www.ciens.ucv.ve/instzool/PropPlan.html.
35. Lentini Z, Martínez CP, Roca WM. Cultivo de anteras de arroz en el desarrollo de germoplasma. Cali: Centro Internacional de Agricultura Tropical; 1997.
36. Borrero J, Martínez CP, Bruzzone C. Introgresión del Nuevo Tipo de Planta desarrollado en el IRRI en los Acervos genéticos de América latina y el Caribe. Culoz: CIAT; 2013.
37. Correa-Victoria FJ, Prado G. Estrategias para el desarrollo de resistencia durable del añublo. Cali: Centro Internacional de Agricultura tropical (CIAT); 2004.
38. Pérez-Almeida I, Lentini Z, Guimarães EP. El cultivo de anteras en el desarrollo de germoplasma resistente al añublo del arroz (Pyricularia grisea). [en línea]. Fitopatología Venezolana. 1995;8(1):11-4.
39. Prado GA, Correa FJ, Aricapa MG, Tulande E, Escobar F. Hipótesis de la exclusión de linajes: una alternativa para el desarrollo de cultivares de arroz con resistencia durable a Pyricularia grisea Sacc en Colombia. Fitopatología Colombiana. 1999;23(2):54-8.
40. Jia Y, Wang Z, Fjellstrom RG, Moldenhauer KA, Azam MA, Correll J, et al. Rice Pi-ta gene Confers Resistance to the Major Pathotypes of the Rice Blast Fungus in the United States. Phytopathology. 2004;94(3):296-301.
41. Jia Y. Artificial introgression of a large chromosome fragment around the rice blast resistance gene Pi-ta in backcross progeny and several elite rice cultivars. Heredity (Edinb). 2009;103(4):333-9.
42. Séré Y, Onasanya A, Afolabi A, Mignouna HD, Akator K. Genetic diversity of the blast fungus, Magnaporthe grisea (Hebert) Barr, in Burkina Faso. Afr J Biotechnol. 2007;6(22):2568-77.
43. Infoagro. La Piricularia del arroz. Corrientes: Universidad Nacional del Nordeste. 2015 [cited 2015 Jun 15]. Available from: http://www.infoagro.com/herbaceos/cereales/arroz.htm.
44. McNally KL, Bruskiewich R, Mackill D, Buell CR, Leach JE, Leung H. Sequencing multiple and diverse rice varieties. Connecting whole-genome variation with phenotypes. Plant Physiol. 2006;141(1):26-31.
45. Hernández J. Mejoramiento para resistencia a plagas con énfasis en Tagosodes orizicolus, Piricularia grisea y Steneotarsonemus spinki. Sancti Spíritus: Proyecto Regional FAO TCP/RLA 3102; 2006.
46. Fuentes JL, Correa-Victoria FJ, Escobar F, Prado G, Aricapa G, Duque MC, et al. Identification of microsatellite markers linked to the blast resistance gene Pi-1(t) in rice. Euphytica. 2008;160(3):29.
Received in May, 2016.
Accepted in November, 2016.
Noraida de Jesús Pérez León. Unidad Científico Tecnológica de Base (UCTB) Los Palacios, Instituto Nacional de Ciencias Agrícolas, INCA. Carretera a Tapaste, Km. 3 1/2 San José de las Lajas, CP 32700, Mayabeque, Cuba. E-mail: nory@inca.edu.cu.


{kind=link}
{kind=link}