










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkBiotecnología Aplicada
versión On-line ISSN 1027-2852
Biotecnol Apl vol.34 no.2 La Habana abr.-jun. 2017
RESEARCH
Generation of a soluble and active recombinant human secretory leukocyte protease inhibitor
Obtención de un inhibidor de proteasas de leucocitos secretores humanos recombinante soluble y activo
Elly Munadziroh1, Shindy Purnamasari2, Ni Nyoman Tri Puspaningsih2, Soetjipto3, Muhammad Rubianto4, Wangsa Tirta Ismaya5*
1 Department of Dental Material and Technology, Faculty of Dentistry, Airlangga University, Mayjen Prof. Dr. Moestopo 47, Surabaya 60132, Indonesia.
2 Department of Biochemistry, Faculty of Science and Technology, Airlangga University,Kampus C UNAIR Mulyorejo, Surabaya 60115, Indonesia.
3 Department of Biochemistry, Faculty of Medicine, Airlangga University, Mayjen Prof. Dr. Moestopo 47, Surabaya 60132, Indonesia.
4 Department of Periodontic, Faculty of Dentistry, Airlangga University Mayjen Prof. Dr. Moestopo 47, Surabaya 60132, Indonesia.
5 Department of Chemistry, Faculty of Natural Sciences, Padjadjaran University, Singaperbangsa 2, Bandung 40133, Indonesia.
*Current address: Dexa Laboratories of Biomolecular Sciences, Industri Selatan V PP-7, Jababeka II Industrial Estate, Cikarang 17550, Indonesia.
ABSTRACT
Inhibition of elastase and matrix metalloprotease activities accelerates the wound healing process. Human secretory leukocyte protease inhibitor (hSLPI) has been reported to demonstrate such inhibition, promoting its use as biomaterial in protein therapy. However, bioavailability of SLPI is very low and its heterologous production in Escherichia coli results in the formation of inclusion bodies, requiring refolding steps to recover its bioactivity. Current strategies to produce SLPI are hampered by either low recovery or process related limitations. Therefore, an alternative approach was developed in order to achieve soluble expression of the protein in E. coli. The gene coding for full-length human SLPI was generated from amniotic membrane through reverse-transcript PCR, and it was then cloned into the pET-101/D-TOPO vector for its expression in E. coli. Soluble recombinant hSLPI with C-terminus His tag was expressed and purified in a nickel affinity column. The purified enzyme was tested for inhibition of elastase. Soluble expression of hSLPI was achieved with the full-length protein containing its native signal peptide. Inhibition of elastase activity by the purified protein indicated that the protein is active, with an inhibition constant of ~9 × 10-8 M, which is in similar order of magnitude to the partial hSLPI. The presence of the signal peptide appears to contribute for the soluble expression of the protein and had negligible effect on its activity.
Keywords: Elastase inhibition, Escherichia coli, native signal peptide, secretory leukocyte protease inhibitor.
RESUMEN
La inhibición de las actividades de elastasa y metaloproteasa de matriz acelera el proceso de cicatrización de heridas. Se ha informado que el inhibidor de la proteasa de leucocitos secretores humanos (hSLPI) demuestra tal inhibición, promoviendo su uso como biomaterial en la terapia de proteínas. Sin embargo, la biodisponibilidad de SLPI es muy baja y su producción heteróloga en Escherichia coli da lugar a la formación de cuerpos de inclusión, requiriendo etapas de replegamiento para recuperar su bioactividad. La estrategia actual para producir SLPI se ve obstaculizada por la baja recuperación o limitaciones relacionadas con el proceso. Por lo tanto, se desarrolló un enfoque alternativo para conseguir la expresión soluble de la proteína en E. coli. Gene codificación de toda la longitud humana SLPI se generó a partir de la membrana amniótica a través de transcripción inversa PCR. El gen obtenido se insertó entonces en pET-101 / D-TOPO para la expresión en E. coli. La hSLPI recombinante soluble con Histag terminal C se expresó y purificó en una columna de afinidad de níquel. Se ensayó la enzima purificada para la inhibición de la elastasa. La expresión soluble de hSLPI se consiguió con la proteína de longitud completa que contenía su péptido señal nativo. La inhibición de la actividad de la elastasa por la proteína purificada sugiere que la proteína es activa, con una constante de inhibición de ~ 9 x 10-8 M, que está en orden similar de magnitud a la hSLPI parcial. La presencia del péptido señal parece contribuir a la expresión soluble de la proteína y parece tener un efecto insignificante sobre la actividad.
Palabras clave: inhibición de elastasa, Escherichia coli, péptido señal nativo, inhibidor de la proteasa de leucocitos secretores.
INTRODUCTION
Human secretory leukocyte protease inhibitor (SLPI) is a non-glycosylated protein that demonstrates anti-protease, anti-inflammatory, and anti-microbial activities [1]. SLPI occurs ubiquitously in human body fluids including tears, milk, saliva, cervical and bronchial mucus, nasal fluid and seminal plasma [2, 3]. The protein plays an important role in the protection of epithelial tissues from serine proteases, such as chymotrypsin and trypsin, leukocyte and neutrophile elastase, and cathepsin G [2], which mostly are involved in degradation of tissues. Therefore, SLPI is often associated with wound healing [4]. SLPI also suppresses the activation of the inflammatory response nuclear factor kB (NF-kB), which prevents stimulation of the release of cytokines (such as interleukin-1β and interleukin-10) and prevents the increase of matrix metalloprotease (MMP) [4]. The aforementioned activities protect epithelial tissue from destruction and allow its recovery. .
Additionally, SLPI decreases the production of interleukin-8 in macrophages upon induction by bacterial lipopolysaccharide (LPS) [5] and prevents the interaction between the human immunodeficiency virus-1 (HIV-1) and human macrophage cells [6]. Furthermore, during herpes simplex virus infection, the expression of SLPI is down regulated [7]. These latter reports indicated a link between SLPI and viral infection. Recently, SLPI has also being associated with anti-tumor activity because its expression results in a blockage of protease-dependent tumor cell migration [8]. Thus, SLPI is an attractive and potential therapeutic protein candidate.
Based on its natural properties, SLPI can be employed to accelerate the healing of gingival injury caused by traction or surgery upon the treatment of gingival recession [9]. Recovery of the wounded gum tissues is often hampered by difficulties to keep the oral cavity free from bacterial infection [10] that may lead to cellulites, abscess and osteomyelitis. Currently, the wound caused by the gingival recession is usually protected by periodontal pack without further treatment to accelerate the healing process or to prevent infection. In this sense, SLPI can be recruited as a biomaterial to accelerate the healing process, as an additional therapy to the current post gingival recession treatment.
One of the sources for the obtainment of SLPI is the amniotic membrane, due to the role of this molecule as defensive factor during pregnancy [3]. In fact, amniotic membrane has successfully been applied to accelerate the healing process of the wound caused by burns [12]. SLPI from amniotic membrane has also been tested in opthalmopathy [13]. Unfortunately, bioavailability of human SLPI (hSLPI) is very low [14] and its commercial recombinant preparation (rhSLPI) (R&D systems, Minneapolis, MD, USA) is expensive (250 US$ for 100 mg). The commercial rhSLPI consists of the mature protein (lacking the signal peptide) that is produced in the bacterial Escherichia coli system as insoluble aggregate so called inclusion bodies, requiring a refolding procedure to recover its bioactivity [15]. This commercial rhSLPI has successfully been tested for therapeutic applications, particularly in accelerating wound healing [16].
Other attempts to produce affordable rhSLPI have achieved limited success. For instance, the use of insect cells is hampered by high cost and poor yields [17]. In the methylotropic yeast Pichia pastoris, reasonable amounts of rhSLPI have been obtained [14] but its production is time consuming (3-4 days) and requires the use of toxic methanol as inducer, which has to regularly be added during production [18]. Moreover, this yeast-derived rhSLPI is likely glycosylated [14], creating a reservation for its future use in protein-based therapy. Further, an expression study of SLPI using S. cerevisiae yeast as the host suggests that the type of signal peptide fused is important for obtaining soluble and active expression of the protein [19]. The expression of both, the mature and truncated (only the C terminal domain) protein species in the presence of invertase as signal peptide results in mixture of active and inactive rhSLPI [19]. The inactive rhSLPI results from the imperfect processing of the signal peptide [19], and other alternatives of signal peptides as that from the α-factor has proven less successful [19]. Hence, this situation may hamper the large scale purification process and may lead to low recovery of the protein. Consequently, these problems to produce SLPI as a fully active preparation and devoid of immunogenicity issues have limited the availability of an SLPI formulation for therapeutic applications.
Alternatively, the production of recombinant proteins employing E. coli is still considered as the best alternative to provide higher yields. This bacterial expression system may also overcome the immunogenicity issue due to the lack of ability of the bacterial system to perform post-translational modifications, while providing soluble and biologically active recombinant hSLPI. The strategy reduces the burden to perform refolding, which is the critical and problematic step to recover biological activity upon large-scale production. At the same time, the expression of hSLPI in S. cerevisiae suggests a possible effect of the signal peptide on protein solubility, despite this peptide is not functional in bacteria. However, the possibility to obtain soluble and biologically active hSLPI in the presence of its native signal peptide (rfhSLPI) in the E. coli system has never been tested. Therefore, in this work, a construct was generated coding for the hSLPI with its native signal peptide. The protein was expressed in E. coli and its activity studied.
MATERIALS AND METHODS
Chemicals and bacterial strains
All chemicals were purchased from Sigma (St. Louis, MO, USA) or Merck (Darmstadt, Germany) except when specifically mentioned. E. coli BL21 star (DE3) and TOP10 strains, pET101D-TOPO cloning vector were purchased from Invitrogen (Carlsbad, CA, USA). The restriction enzymes were from MBI Fermentas (Hannover, MD, USA). The DNA plasmid was isolated and purified using Miniprep DNA extraction kit from Qiagen (Chatsworth, CA, USA). The porcine pancreatic elastase (PPE) and the N-succinyl- L-alanyl-L-alanyl-L-prolyl-L-phenylalanyl-4-nitro-analide substrate were kindly provided by Prof. Kim Kyung-Min of Kyungpook National University, Daegu, South Korea. The ProfinityTM IMAC Nickel-IDA resin was purchased from Bio-Rad (Singapore).
Generation of rhSLPI
The rhSLPI encoding gene was inserted to pET- 101/D-TOPO and cloned into BL21 Star (DE3). A 2-L Erlenmeyer flask containing 400 mL of LB medium supplemented with 100 μg/mL ampicillin was inoculated by picking a single E. coli colony from an LB plus ampicillin (100 μg/mL) agar plate, and incubated at 37 ºC for 18 h under constant agitation (150 rpm). Subsequently, the culture was inoculated into the production culture medium of the same compo-sition (1 %, v/v) and it was further grown at 37 ºC under agitation at 150 rpm. The expression of rhSLPI was induced when the OD600 nm reached 0.5-0.8 by the addition of isopropyl thiogalactopyranoside (IPTG) at concentrations below 50 μM. After 4 h of culture, bacterial cells were harvested by centrifugation at 5500 rpm and 4 ºC for 10 min.
Then, cells were washed with 50 mM phosphate buffer, pH 8.0, containing 300 mM NaCl, and resuspended in phosphate buffer (volume). The cell suspension was then sonicated seven times for 15 s at 30 s intervals. The cell free extract was collected by centrifugation at 5500 rpm at 4 ºC for 15 min. Proteins in the supernatant fraction were then concentrated by means of acetone precipitation method [20]. The precipitated proteins were dissolved in the same buffer supplemented with 10 mM imidazole. After incubation at 4 ºC with the Nickel-IDA affinity resin, the rhSLPI was eluted from the affinity matrix in a column with 50 mM phosphate buffer, pH 8.0, containing 300 mM NaCl and 175 mM imidazole.
Analysis of purified rhSLPI
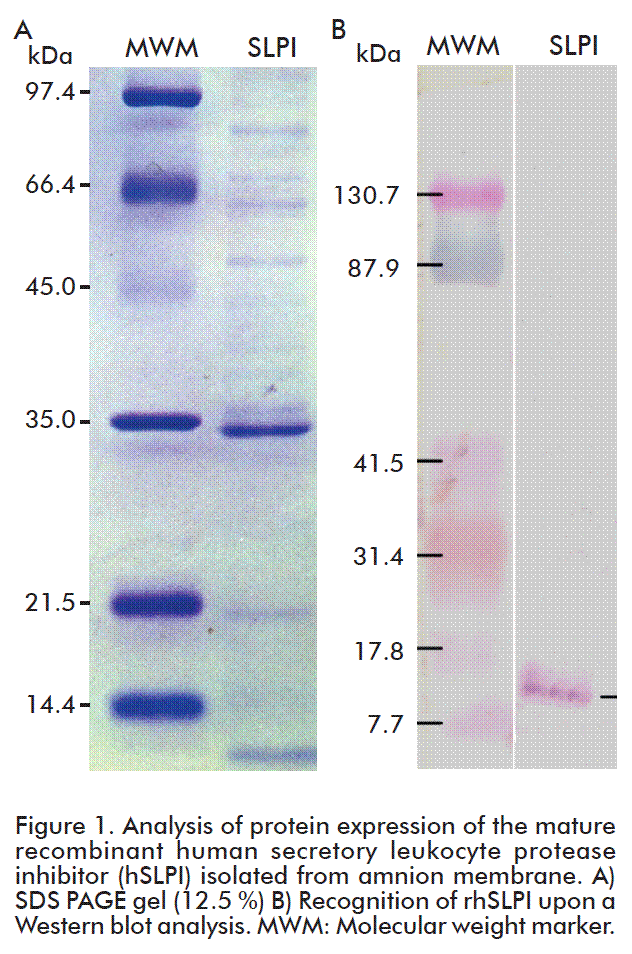
The purity of rhSLPI after one step purification on an affinity chromatography was evaluated by SDS PAGE analysis (12.5 %). The identity of rhSLPI was confirmed by western blot analysis using monoclonal antibody raised against hSLPI, and its concentration was determined by the Bradford’s method using BSA as standard. The western blot analysis was performed using monoclonal mouse antibody raised against full-length hSLPI (Santa Cruz Biotechnology, Dallas, TX, USA), visualized with conjugate anti-mouse alkaline phosphatase (Promega, Madison, WI, USA).
Activity assay of SLPI using PPE
Briefly, into a cuvette in a spectrometer with temperature controlled at 25-30 ºC, 750 μL substrate N-succinyl-L-alanyl-L-alanyl-L-prolyl-L-phenylalanyl-4-nitroanalide (NPN) was added to (750 – X) μL of 100 mM HEPES buffer, pH 7.4, , where X is the volume of rhSLPI added). Immediately after the addition of rhSLPI, the release of p-nitroaniline (PNA) was followed by detection at 410 nm for 4 min every 30 s. The NPN substrate concentrations tested ranged 1-3 μM (using incremental steps of 0.5 μM). The activity of PPE in the absence of SLPI was determined accordingly by replacing SLPI with buffer.
RESULTS AND DISCUSSION
Expression of soluble rfhSLPI in E. coli BL21 Star (DE3) was achieved upon induction at IPTG concentrations of 50 μM or less. An SDS PAGE analysis after one step of purification on Ni-NTA affinity column suggested the presence of the soluble rfhSLPI that was confirmed with western blot analysis (Figure 1). The size of the expected protein band appeared to agree with the rfhSLPI gene construct that contained the native signal peptide and poly-histidine tag (His-tag). Secondary structure prediction of the signal peptide region suggested occurrence of α-helical structure (15 residues long) [21]. This helical structure may alter the structure of the mature hSLPI (residues 26-132), which contains only little secondary structures that consist of two-short β-strands (of four residues each) and a short turn α-helix (also of four residues) with no hydrophobic core [22]. The presence of the signal peptide has probably increased the solubility of rfhSLPI. This finding may concur with the reported study on overexpression of both the full-length and mature hSLPI in keratinocytes that suggests crucial functioning of the signal peptide for secretion of the protein out from the cells, for prevention against intracellular degradation, and also for its synthesis [23].
Biological activity of the soluble rfhSLPI was evaluated from its capability to demonstrate inhibition to the elastase activity, which is based on the amout of p-nitroaniline (PNA) released during hydrolysis of the NPN substrate. In the presence of 9.25 μg rfhSLPI, the PPE activity was decreased from 16.48 ± 0.10 U/mg to 14.03 ± 0.28 U/mg. SLPI has indeed been reported to display weaker inhibition to PPE than human neutrophil elastase (HNE) [24]. Inhibition of the elastase activity is originated from interactions involving only the C-terminal region of SLPI [25]. In detailed, the crystal structure of PPE complex with the C terminal domain of hSLPI (PDB ID 2Z7F) shows interactions between the active site of PPE (residues Phe41, Gly193, Ser195, Ser217-Val216-Phe215-Ser214) and a surface loop in the C terminal region of hSLPI (residues Gly69-Gln70-Cys71- Leu72-Met73-Leu74) [25]. Based on this structural information, hSLPI is categorized as the canonical serine protease type of inhibitor that binds the enzyme target through an exposed and convex binding loop complementary to the active site [26]. Although the inhibition profile appears to be weak (~15 %), specificity of hSLPI interactions with PPE suggests that the observed PPE activity inhibition may not be artificial. Furthermore, the presence of the signal peptide and the C terminal His-tag appear not to abolish rfhSLPI function to inhibit elastase activity.
Additional kinetics analysis (Figure 2) indicated that rfhSLPI appears to display a mix mode inhibition, as both substrate binding and rate of catalysis were affected. The inhibition constant (Ki) of the full-length rhSLPI in this study was ~ 9 × 10-8 M, similar to that of the commercial preparation (~5 × 10-8 M). The result of this study falls in the range of the reported Ki value for the C domain rhSLPI (at ≥ 50 × 10-9 M) [27]. In a mix mode of inhibition, elastase could be inhibited through interactions between SLPI with the active site of the enzyme or somewhere on the enzyme molecule that interferes with the enzyme-substrate interaction. This phenomenon seems unlikely because interaction between hSLPI and PEE is rather specific. However, if the lowest substrate concentration is excluded from the data, the inhibition profile appears to resemble more of the uncompetitive type, which means rfhSLPI binds only to elastase-substrate complex. This latter hypothesis on the inhibition mode is more likely. At the lowest substrate concentration, the inhibition may actually be insignificant because of the Ki value of SLPI for PPE is rather high in comparison to other elastase inhibitors [27]. Nevertheless, the activity assay and kinetics evaluation show that the full-length rhSLPI obtained is biologically active.
The presence of the native signal peptide appears to be harmless for the inhibitory activity of SLPI. Previously, the swapping of the N and C terminal domains of SLPI or fusion of some protease inhibitory proteins to the C terminal domain is reported to cause changes in the Ki value [28]. Unfortunately these studies did not report the mode of inhibition of those other rhSLPI variants. Nevertheless, the difference in the Ki value of rfhSLPI was anticipated.
Our study demonstrates the possible use of E. coli system for expression of soluble SLPI in the presence of its native signal peptide. The helical structure of the signal peptide may be altering the structure of mature SLPI, which contains only little secondary structures. Furthermore, the rfhSLPI appeared to remain biologically active in the presence of its native signal peptide and a C terminal His-tag. The two additional entities seem to do not interfere with the functioning of rfhSLPI. However, their potential immunogenicity should be evaluated prior to its development as therapeutic agent. While the His-tag is considered to have low level of toxicity and immunogenicity [29], the presence of the signal peptide is not yet known. Thus, while the C terminal His-tag could be left intact, removal of the signal peptide may require a strategy, since SLPI inhibits the activity of proteases that are employed in protein maturation [25].
Moreover, the use of a synthetic gene with optimized codon usage increased the expression level but the protein synthesis rate must be kept at minimum as suggested in this study, in which soluble rhSLPI was obtained only at low IPTG concentrations (less than 50 μM). Targeting rhSLPI to the periplasmic space could be an attractive alternative as the presence of DsbA/DsbB chaperone system that assists the formation of disulfide bonds may also overcome problems for a proper protein folding. Participation of protein disulfide isomerase (PDI) that facilitates the formation of disulfide bonds during the expression of rhSLPI in the yeast Pichia pastoris, increases the yield of the recombinant protein up to 5-8 folds [30]. Additionally, targeting to periplasmic space might cause another problem with the yield because the periplasmic space can accommodate only for less than 10 % of the total protein content of the E. coli cells. As for expression in P. pastoris, the issue with potential immunogenicity is not yet resolved. Thus, bacterial cytoplasmic production of SLPI in its soluble form is still the most appealing alternative. Our report suggested that production of soluble and biologically active rfhSLPI in E. coli was made possible through the inclusion of its native signal peptide. This approach appears to be a promising alternative for production of recombinant SLPI. Our next target is to improve the level of recombinant protein expression without compromising its solubility.
REFERENCES
1. Doumas S, Kolokotronis A, Stefanopoulos P. Anti-inflammatory and antimicrobial roles of secretory leukocyte protease inhibitor. Infect Immun. 2005;73(3):1271-4.
2. Moreau T, Baranger K, Dade S, Dallet- Choisy S, Guyot N, Zani ML. Multifaceted roles of human elafin and secretory leukocyte proteinase inhibitor (SLPI), two serine protease inhibitors of the chelonianin family. Biochimie. 2008;90(2):284-95.
3. Zhang Q, Shimoya K, Moriyama A, Yamanaka K, Nakajima A, Nobunaga T, et al. Production of secretory leukocyte protease inhibitor by human amniotic membranes and regulation of its concentration in amniotic fluid. Mol Hum Reprod. 2001;7(6):573-9.
4. Ashcroft GS, Lei K, Jin W, Longenecker G, Kulkarni AB, Greenwell-Wild T, et al. Secretory leukocyte protease inhibitor mediates non-redundant functions necessary for normal wound healing. Nat Med. 2000;6(10):1147-53.
5. Weldon S, McGarry N, Taggart CC, McElvaney NG. The role of secretory leucoprotease inhibitor in the resolution of inflammatory responses. Biochem Soc Trans. 2007;35(Pt 2):273-6.
6. Kazmi SH, Naglik JR, Sweet SP, Evans RW, O’Shea S, Banatvala JE, et al. Comparison of human immunodeficiency virus type 1-specific inhibitory activities in saliva and other human mucosal fluids. Clin Vaccine Immunol. 2006;13(10):1111-8.
7. Fakioglu E, Wilson SS, Mesquita PM, Hazrati E, Cheshenko N, Blaho JA, et al. Herpes simplex virus downregulates secretory leukocyte protease inhibitor: a novel immune evasion mechanism. J Virol. 2008;82(19):9337-44.
8. Wen J, Nikitakis NG, Chaisuparat R, Greenwell-Wild T, Gliozzi M, Jin W, et al. Secretory leukocyte protease inhibitor (SLPI) expression and tumor invasion in oral squamous cell carcinoma. Am J Pathol. 2011;178(6):2866-78.
9. Newman MG, Takei H, Klokkenvold PR, Carranza FA. Carranza’s Clinical Periodontology. 11th Ed. Philadelphia: W.B. Saunders Co.; 2011.
10. Kassab MM, Cohen RE. The etiology and prevalence of gingival recession. J Am Dent Assoc. 2003;134(2):220-5.
11. Moriyama A, Shimoya K, Ogata I, Kimura T, Nakamura T, Wada H, et al. Secretory leukocyte protease inhibitor (SLPI) concentrations in cervical mucus of women with normal menstrual cycle. Mol Hum Reprod. 1999;5(7):656-61.
12. Ley-Chavez E, Martinez-Pardo ME, Roman R, Oliveros-Lozano Fde J, Canchola-Martinez E. Application of biological dressings from radiosterilized amnios with cobalt 60 and serologic studies on the handling of burns in pediatric patients. Ann Transplant. 2003;8(4):46-9.
13. Dua HS, Gomes JA, King AJ, Maharajan VS. The amniotic membrane in ophthalmology. Surv Ophthalmol. 2004;49(1):51-77.
14. Li Z, Moy A, Sohal K, Dam C, Kuo P, Whittaker J, et al. Expression and characterization of recombinant human secretory leukocyte protease inhibitor (SLPI) protein from Pichia pastoris. Protein Expr Purif. 2009;67(2):175-81.
15. Kikuchi T, Abe T, Yaekashiwa M, Tominaga Y, Mitsuhashi H, Satoh K, et al. Secretory leukoprotease inhibitor augments hepatocyte growth factor production in human lung fibroblasts. Am J Respir Cell Mol Biol. 2000;23(3):364-70.
16. Nakamura K, Takamoto N, Hongo A, Kodama J, Abrzua F, Nasu Y, et al. Secretory leukoprotease inhibitor inhibits cell growth through apoptotic pathway on ovarian cancer. Oncol Rep. 2008;19(5):1085-91.
17. Gray LR, Alexander AL, Shugars DC. Construction, non-denaturing affinity purification, and characterization of baculovirally expressed human secretory leukocyte protease inhibitor. Protein Expr Purif. 2002;26(1):179-86.
18. Cereghino JL, Cregg JM. Heterologous protein expression in the methylotrophic yeast Pichia pastoris. FEMS Microbiol Rev. 2000;24(1):45-66.
19. Stetler GL, Forsyth C, Gleason T, Wilson J, Thompson RC. Secretion of active, full-and half-length human secretory leukocyte protease inhibitor by Saccharomyces cerevisiae. Nat Biotechnol. 1989;7:55-60. .
20. Simpson DM, Beynon RJ. Acetone precipitation of proteins and the modification of peptides. J Proteom Res. 2010;9(1):444-50.
21. Buchan DW, Ward SM, Lobley AE, Nugent TC, Bryson K, Jones DT. Protein annotation and modelling servers at University College London. Nucleic Acids Res. 2010;38(Web Server issue):W563-8.
22. Lin CC, Lu BY, Chang JY. Conformational stability of secretory leucocyte protease inhibitor: a protein with no hydrophobic core and very little second-ary structure. Biochim Biophys Acta. 2006;1764(7):1286-91.
23. Jacobsen LC, Sorensen OE, Cowland JB, Borregaard N, Theilgaard-Monch K. The secretory leukocyte protease inhibitor (SLPI) and the secondary granule protein lactoferrin are synthesized in myelocytes, colocalize in subcellular fractions of neutrophils, and are coreleased by activated neutrophils. J Leukoc Biol. 2008;83(5):1155-64.
24. Masuda K, Suga T, Takeuchi A, Kanesaki M, Imaizumi A, Suzuki Y. Specific cleavage of secretory leukoprotease inhibitor by neutrophil elastase and saliva. Biochem Pharmacol. 1994;48(4):651-7.
25. Koizumi M, Fujino A, Fukushima K, Kamimura T, Takimoto-Kamimura M. Complex of human neutrophil elastase with 1/2SLPI. J Synchrotron Radiat. 2008;15(Pt 3):308-11.
26. Otlewski J, Krowarsch D, Apostoluk W. Protein inhibitors of serine proteinases. Acta Biochim Pol. 1999;46(3):531-65.
27. Zani ML, Baranger K, Guyot N, Dallet-Choisy S, Moreau T. Protease inhibitors derived from elafin and SLPI and engineered to have enhanced specificity towards neutrophil serine proteases. Protein Sci. 2009;18(3):579-94.
28. Zani ML, Nobar SM, Lacour SA, Lemoine S, Boudier C, Bieth JG, et al. Kinetics of the inhibition of neutrophil proteinases by recombinant elafin and pre-elafin (trappin-2) expressed in Pichia pastoris. Eur J Biochem. 2004;271(12):2370-8.
29. Walls D, Loughran ST. Tagging recombinant proteins to enhance solubility and aid purification. Methods Mol Biol. 2011;681:151-75.
30. Li Z, Moy A, Gomez SR, Franz AH, Lin-Cereghino J, Lin-Cereghino GP. An improved method for enhanced production and biological activity of human secretory leukocyte protease inhibitor (SLPI) in Pichia pastoris. Biochem Biophys Res Commun. 2010;402(3):519-24.
Received in October, 2016.
Accepted in February, 2017.
Elly Munadziroh. Department of Dental Material and Technology, Faculty of Dentistry, Airlangga University, Mayjen Prof. Dr. Moestopo 47, Surabaya 60132, Indonesia. E-mail: elly-m@fkg.unair.ac.id, munadziroh.elly@gmail.com.


{kind=link}
{kind=link}