










My SciELO
 Custom services
Custom servicesServices on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkBiotecnología Aplicada
On-line version ISSN 1027-2852
Biotecnol Apl vol.35 no.1 La Habana Jan.-Mar. 2018
RESEARCH
Fermentative behavior of the Candida stellata yeast under different aeration conditions
Comportamiento fermentativo de la levadura Candida stellata en diferentes condiciones de aireación
Waldir D Estela-Escalante1,2, Gloria A Cosco-Salguero1, Elena L Quillama-Polo1, Mojmír Rychtera2
1 Escuela Profesional de Ingeniería Agroindustrial. Facultad de Química e Ingeniería Química, Universidad Nacional Mayor de San Marcos. Av. Universitaria s/n. Lima 1, Lima, Perú.
2 Department of Fermentation Chemistry and Bioengineering. Faculty of Food and Biochemical Technology. University of Chemistry and Technology. Prague 6, Dejvice. Czech Republic.
ABSTRACT
The influence of aeration on the fermentative activity of Candida stellata RIVE 3-16-1 was studied. The strain was cultured either in Erlenmeyer flasks or a bioreactor containing sterilized and aroma-removed apple juice. Cultures in Erlenmeyer flasks were done under shaken or static condition, whereas in the bioreactor, a constant air flow regime was kept. Chemical compounds produced during fermentation were determined by GC and HPLC. The agitation of Erlenmeyer flasks increased the production of total higher alcohols as compared to static culture and enhanced dramatically the ethyl acetate production. Meanwhile, the production of acetic acid and glycerol were higher under static culture. Bioreactor fermentation at constant air flow was important to visualize the effect of oxygen on the production of compounds and its impact in the quality of alcoholic beverages. It is reported a specific growth rate of 0.13 h-1. Aireation promoted the cell growth affecting the ethanol yield. At the end of culture, malic acid naturally present in apple juice and the ethanol produced were consumed after sugar depletion. Moreover, the acetic acid produced at the end of the fermentation served as carbon source too. The best results in terms of acceptability of the fermented beverages were obtained when cultivated statically.
Keywords: Candida stellata, alcoholic fermentation, higher alcohols, ethyl acetate.
RESUMEN
Se evaluó la influencia de la aireación en la actividad fermentativa de Candida stellata RIVE 3-16-1. La cepa se cultivó en frascos de Erlenmeyer y en un biorreactor que contenía jugo de manzana esterilizado y sin aroma. El cultivo en erlenmeyers se realizó en condiciones estáticas o con agitación, mientras queen el biorreactor se mantuvo bajo régimen constante de flujo de aire. Los compuestos químicos producidos durante la fermentación se determinaron por cromatografía de gases y cromatografía líquida de alta presión (HPLC). La agitación de los frascos de Erlenmeyer incrementó la producción de alcoholes superiores en comparación con el cultivo estático y mejoró acentuadamente la producción de etil acetato. Mientras tanto, la producción de ácido acético y glicerol fue mayor en el cultivo estático. El efecto del oxígeno en la producción de compuestos y su impacto en la calidad de bebidas alcohólicas se visualize en cultivo en biorreactor a flujo de aire constante. Se observó una tasa de crecimiento específico de 0.13 h-1. La aireación promovió el crecimiento celular afectando el rendimiento de etanol. Al final del cultivo, el ácido málico presente naturalmente en el jugo de manzana y el etanol producido fueron consumidos después del agotamiento del azúcar. Adicionalmente, el ácido acetico producido al final del proceso sirvió como fuente de carbono. Los mejores resultados en términos de aceptabilidad de las bebidas fermentadas se obtuvieron en cultivo estático.
Palabras clave: Candida stellata, fermentación alcohólica, alcoholes superiores, etil acetato.
INTRODUCTION
Non-Saccharomyces yeasts have been always regarded as contaminants in the production of wines, ciders and beers. For this reason, no attention has been paid to the potential use of these yeasts for fermented beverages production processes. Just few studies have addressed their fermentative behaviour and the factors influencing it, with Kloeckera apiculata, Saccharomycodes ludwigii, Hansenula anomala and Hanseniaspora uvarum as the most relevant non-Saccharomyces yeasts studied in the last years [1-4]. All of them showed that these yeasts are strongly influenced by the presence of oxygen in the fermentation medium and, consequently, it impacts in the production of compounds of sensory importance, such as esters and higher alcohols.
Among the non-Saccharomyces yeasts, Candida spp. are gaining importance from the industrial point of view due to its particular fermentative behavior. For instance, some Candida species are being considered potential candidates for the fermentation of wines and beers respectively [5-7]. Of them, Candida stellata is commonly isolated from grape must and survives during spontaneous wine fermentation for longer periods of time [8, 9]. Studies on the fermentative activity of C. stellata showed that it could positively affect the taste and flavour of alcoholic beverages [10]. Moreover, this yeast shows a strong fructophilic and osmophilic character [11] and it is a potent producer of glycerol under aerobic growth conditions and especially in anaerobic ones [12]. Regarding its metabolism, some authors refer to C. stellata as Crabtree positive yeast [13], since it shows different metabolic behaviour depending on the oxygen and glucose concentration in the fermentation medium. Under anaerobic or oxygenlimited conditions, this type of yeasts exhibit alcoholic fermentation [14], but in fully aerobic ones, a mixed respiro-fermentative metabolism is observed due to the excess of sugar concentration over a given threshold value. That value tends to be about 1 mM in Saccharomyces cerevisiae [15]. Conditions leading to sugar fermentation result in the production of major fermentative compounds in detriment of biomass yield, including ethanol, acetic acid and glycerol, with small amounts of higher alcohols, esters, volatile fatty acids and carbonyl compounds (less than 1 % w/v of the utilized sugar). Due to these metabolic losses, the complete fermentation of hexose by yeasts yields 94-96 % of the theoretical ethanol yield [16].
While complementary to major fermentative products, those minoritary chemical compounds are of paramount sensory importance, including esters, higher alcohols, organic acids, aldehydes and others. In fact, esters are the most important flavour compounds in many alcoholic beverages such as beer and wine [17]. Esthers are produced from the reaction between alcohols and fatty acids, which is catalyzed by the enzyme alcohol acetyl transferase (AATase) [18]. Noteworthy, AATase activity is strongly inhibited by trace amounts of oxygen [19]. Regarding higher alcohols, they are produced either catabolically from the degradation of cell-imported amino acids or anabolically via the biosynthetic route from the carbon source [20]. Oxygen supplementation and temperature increase during fermentation enhance the production of higher alcohols by metabolism activation and, consequently, it promotes the cell growth [21].
In order to elucidate the balance between all these processes, in this work it was studied the effect of aeration on the fermentative metabolism of Candida stellata RIVE 3-16-1, and the production of sensory-relevant compounds, using apple juice as fermentation medium. Two culture conditions were considered, attending to their technological importance: fermentation under highly limited oxygen concentration (static culture) or with moderate oxygen (agitated culture). Additionally, a batch culture in a bioreactor containing juice of a specific apple variety under constant air flow was done to evaluate the metabolic fermentative behavior of C. stellata RIVE 3-16-1.
MATERIALS AND METHODS
Microorganism and maintenance
The Candida stellata RIVE 3-16-1 yeast strain was acquired from the collection of yeasts at the Research Institute of Viniculture and Enology of Bratislava, Slovak Republic, as used in the experiments as further specified. The strain was maintained on malt extract agar at 7 ºC and renewed every three months, until use.
Propagation of inoculum
The inoculum was propagated in 100 mL of sterile apple juice at 28 ºC for 24 h. Flasks were shaken at 200 min-1 in an orbital shaker. Cell biomass was separated by centrifugation (3000 rpm for 10 min) and then washed three times with sterile physiological solution. Fermentation media were inoculated with 1.0 ± 0.1 g of cells based on wet weight [1-4].
Synthesis of compounds of sensory importance
Sensory-relevant compounds were produced by fermentation in concentrated, sterilized and aroma-removed apple juice (Severofrukt a.s, Terezin, Czech Republic). It was reconstituted with sterilized distilled water up to a total sugar concentration of 12.8 % w/v and pH 3.8 [1-4, 22]. Fermentations were done in triplicate, both static and agitated at 28 ºC in 500 mL Erlenmeyer flasks containing 250 mL of medium. Under agitation, flasks were shaken at 200 rpm for 8 d. Static cultures were fermented for 15 d, approximately. Fermentations were regarded as finished when sugar was depleted.
Batch culture in bioreactor
Apple juice of Rubin variety containing 13 % w/v of total sugars and pH 3.8 was utilized in the experiments. Apples were acquired from CZ-fruit, (Prague-CR, Czech Republic), and the juice was extracted by pressing and then placed in 10-L glass containers. Then, juice was pasteurized in a thermostat at 65-70 ºC for 10 h (including the cooling time) to eliminate microflora and the varietal volatile compounds [1-4, 23]. Subsequently, juice was supplemented with 1.2 g/L KH2PO4 and 1.2 g/L (NH4)2SO4 as cell growth promoter compounds [1-4, 24].
Cultures were carried out in a 2-L bioreactor (BIOSTAT, B. Braun International, Germany) containing 1.5 L of apple juice. The equipment was provided with a pH-meter, thermometer, stirrer and an electrode for measurement of dissolved oxygen. The bioreactor was connected to a micro-DCU-300 regulation and measurement unit. The following parameters were kept constant during the entire process: temperature, 18 ºC; stirring frequency, 300 rpm; air flow rate, 25 L/h (0.2 mol O2/h). Cultures were stopped when observing an increase of the dissolved oxygen value to its initial value (100 %).
The inoculum was propagated in 80 mL of synthetic medium containing: 100 g/L glucose; 10 g/L peptone, 1.2 g/L KH2PO4, 1.2 g/L (NH4)2SO4 and 10 g/L yeast extract; pH was adjusted to 3.8 with a 5 % v/v HCl solution. Cells were propagated at 28 ºC for 48 h in an orbital shaker at 150 rpm. Subsequently, the cells were separated by centrifugation (3000 rpm for 10 min), washed with sterile physiological solution and finally inoculated into the bioreactor.
Analytical methods
Volatile compounds (higher alcohols and esters) produced during fermentation were analyzed by a gas chromatograph (Hewlett-Packard 5890II), equipped with a HP5 column (30 m × 0.32 mm) and FID detector. The working parameters were: injector temperature, 240 °C; oven conditions, 40 °C initial temperature for 5 min, then was raised to 220 °C at 4 °C/min and kept at this temperature for 5 min. The samples (three replicates for each experiment) were previously centrifuged and then filtered through 0.45 μm micromembranes. Volatile compounds were analyzed using the method of dichloromethane micro extraction [25]. Finally, 1 μL of each extract was injected into the column of the equipment.
Acetic, succinic and malic acids, ethanol, glycerol, fructose and glucose were analyzed by a HPLC (Pump LCP 4000), equipped with a repacking watrex 250 × 8 mm column (Ostion LGKS 0800 H+ form) and a RID detector. Malic acid was analyzed only in fermentations carried out with Rubin apple juice. The conditions of analysis were: column temperature 80 °C, mobile phase 5 mM H2SO4, 1 mL/min flow rate.
Once centrifuged at 10 000 rpm and filtered, samples were diluted with demineralized water (1:3) before injecting to the equipment [1-4, 24].
Cell biomass was determined by gravimetry. Cells were separated by centrifugation (3000 rpm for 10 min), then washed three times with distilled water, dried at 110 °C until constant weight and finally weighed. Additionally, biomass yield and ethanol (YX/S and YE/S) and, the specific growth rate (μ) were determined at the end of the experiment.
Sensory and statistical analyses
Sensory assessment of ciders fermented statically and in agitation was performed using an affective and descriptive analysis in the same manner as in previous studies carried by Estela-Escalante et al. [1-4, 24]. Attributes such as taste, aroma and odor were evaluated using a 5-points Hedonic scale (1, dislike very much; 2, dislike somewhat; 3, neither like nor dislike; 4, like somewhat; 5, like very much). Samples were evaluated by a trained panel of 10 judges, (all men, and aged 25-30 years). The sensory evaluation was done according to Meilgaard et al. [26], and sensory evaluation data were presented as means of the score of all the judges. A standard Student’s t test was used to analyze the statistical significance (p < 0.05) of the differences observed between the scores for two fermented beverages (cultivated under agitation or static). Statistical analysis was done using Statistica v.8.0 software.
RESULTS AND DISCUSSION
Culture under static and agitated conditions
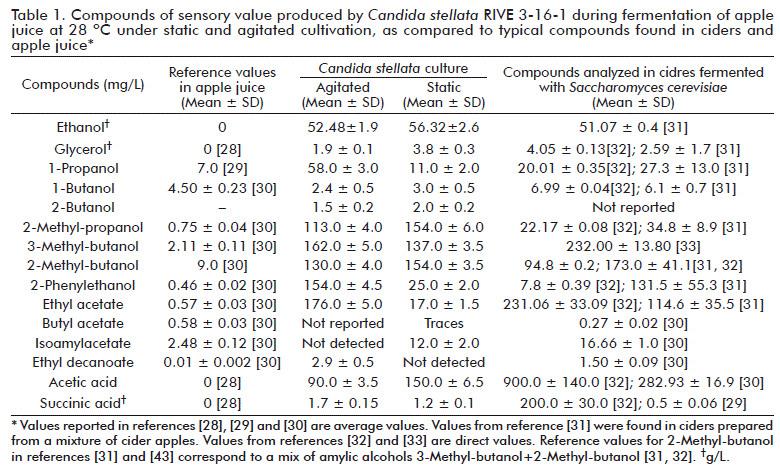
Culture experiments done in shake-flasks are regarded as oxygen-limited [27], this culture condition affecting the fermentative metabolism of yeasts. In fact, high concentration of sugar present in apple juice leads to alcoholic fermentation in Crabtree positive species even when oxygen is present in enough concentration. Culture without agitation (statically) are even more oxygen-limited, since only the surface of the liquid gets into contact with the air and the movement of cells in the liquid is very slow. Oxygen is necessary to improve yeast metabolism and, thus, to successfully complete the fermentation. Results consistent with these observations and derived from the production of chemical compounds during fermentation of apple juice by C. stellata RIVE 3-16-1 in static and agitated cultures are shown in Table 1. Compounds analyzed in ciders fermented with different strains of S. cerevisiae as reported by different authors were included for comparison.
Glycerol production
The production of glycerol under static culture reached 1.9 ± 0.1 g/L, significantly different (p < 0.05; determinations in triplicates) from results in static culture (3.8 ± 0.3 g/L). The decrease of glycerol production under agitated conditions was also observed in previous studies with Kloeckera apiculata, S. cerevisiae, Saccharomycodes ludwigii and Hanseniaspora uvarum under similar culture conditions [1, 2, 4, 24]. Aerobic conditions promote cellular respiration and severely decrease glycerol production. Noteworthy, C. stellata is considered as a high producer of glycerol and amounts up to 11.76 g/L have been reported [34]. Despite, the 3.8 ± 0.3 g/L glycerol obtained in this study is low as compared to values reported in the literature, what would suggest that glycerol production by C. stellata could be strain-dependent. From the technological point of view, a higher production of glycerol is desirable since it influences positively the sensory quality of alcoholic beverages, while not detrimental for alcohol production. Glycerol imparts a slightly sweet taste and contributes to the mouth-feel and complexity of wine flavour at low levels [35, 36].
Production of higher alcohols
Higher alcohols need to be kept low since they are mostly detrimental, with few exceptions, to the sensory quality of fermented beverages. In our experiments, a higher production of total higher alcohols in agitated culture (620.9 mg/L) was observed, in comparison with the static culture (486.0 mg/L). This behavior has been also described for non-Saccharomyces yeasts (K. apiculata, H. uvarum and S. ludwigii) under similar culture conditions [1, 2, 4]. Nevertheless, an opposite effect was evidenced with S. cerevisiae [24]. Hence, considering the present results together with previous ones from our group [1-4, 24], it is possible to suggest that the production of higher alcohols depends on yeast type and culture conditions. From the sensory point of view, higher alcohols are identified by a strong, pungent smell and taste [37, 38]. For instance, total concentrations higher than 400 mg/L negatively contribute to the organoleptic quality of wines [39]. Except for 2-phenylethanol, which imparts a floral aroma [40], the rest of higher alcohols provide unpleasant sensory characteristics [39, 40].
Production of esters
Esters are the most important aroma compounds in alcoholic beverages. They confer pleasant sensory characteristics under certain concentrations. Among them, ethyl acetate is present in most fermented beverages in higher amounts as compared to the rest of acetate esters. It imparts a light-fruity or solvent-like aroma depending on its concentration. Mean values of experiments carried out in triplicate showed that agitation enhanced the production of ethyl acetate (176.0 ± 5.0 mg/L), ten times higher than in static culture (17.0 ± 1.5 mg/L, p < 0.05). These results are opposite to those by Yoshioka and Hashimoto [19], reporting that small amounts of oxygen had inhibitory effects on AATase, the enzyme responsible for the ester generation. Previous results from our group with other non-Saccharomyces yeasts such as K. apiculate and H. uvarum showed that ethyl acetate production diminished as consequence of agitation [1, 4]. On the other hand, in studies with S. cerevisiae, S. ludwiggi and Hansenula anomala carried out under similar cultivation conditions an opposite effect was evidenced [2, 3, 24]. From the sensory point of view, ethyl acetate concentrations lower than 80.0 mg/L contribute positively to the flavour and taste of wines [41]. On the contrary, concentrations over 200.0 mg/L would impart a vinegar taste [42].
Production of organic acids
Two relevant organic acids were determined: acetic acid and succinic acid. Acetic acid is quantitatively and sensorially the most important volatile fatty acid produced during alcoholic fermentation. Yeasts produce different amount of acetic acid during ethanol production. In this study, higher amounts were produced in static (150.0 ± 6.5 mg/L) vs agitated (90.0 ± 3.5 mg/L) conditions. This behavior matched that observed in previous studies with S. cerevisiae and K. apiculata under similar culture conditions [1, 24]. The flavour threshold for acetic acid depends on the type of alcoholic beverage, ranging 0.4-1.1 g/L for wines [43].
In the case of succinic acid, it is the major organic acid produced by yeast metabolism [44], and it has an unusual salty and bitter taste. Metabolically, it is an intermediary compound of the Krebs cycle. No significant differences were found (p > 0.05) of succinic acid production between static (1.2 ± 0.1 g/L) and agitated (1.7 ± 0.15 g/L) conditions, as mean values from three replicates of each fermentation condition. This can be explained due to the fact that agitation oxygenates the medium, promoting the synthesis of this acid.
Sensory evaluation of fermentation product
Significant differences (p < 0.05) were found for the two fermentation beverages (static vs agitated) when comparing the mean values of the score of 10 judges who evaluated them attending to flavour, taste and odour. Panelists described the beverage fermented statically with a slightly taste of solvent-like, little acidic, with an acceptable balanced sensory profile. On the other hand, they referred to the beverage fermented under agitation as of strong taste to solvent like and with unbalanced sensory profile. The beverage fermented statically had an overall acceptability, based on the attributes evaluated, which is consistent with the traditional fermentation process.
Batch cultivation in bioreactor under constant air flow
Batch cultivation with constant air flow regime allows constant supply of oxygen to the medium. Oxygen is the most important parameter determining the balance between fermentative and respiratory activity in most yeasts. Respiration rate becomes dependent on oxygen availability in the medium at concentrations lower than the critical value, this parameter very low for yeasts at about 0.12 mg/L at 20 ºC [45]. Oxygen transfer into the liquid phase depends on several parameters, such as: the bioreactor geometry, type of impeller, culture medium viscosity, agitation speed, number of baffles, temperature, among others. Only the oxygen dissolved in the liquid phase is available for yeast cells.
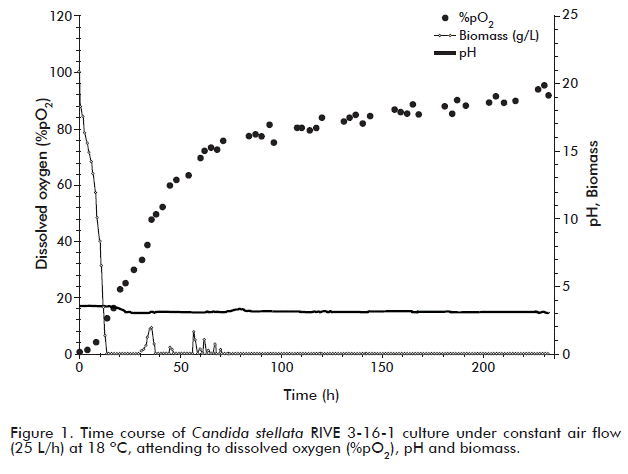
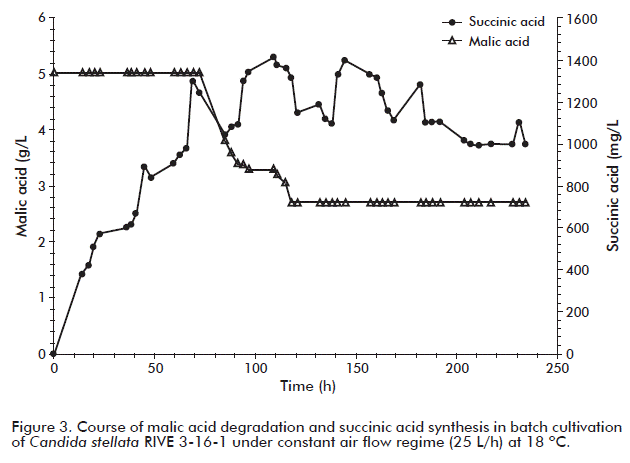
Therefore, the aim was to evaluate the impact of oxygenation on the fermentative behavior of C. stellate RIVE 3-16-1 yeast. For that purpose, apple juice from the single Rubin variety was used and following the routine procedure of juice extraction. Oxygen consumption, cell growth, pH shift and the synthesis and utilization of metabolism byproducts were monitored at a constant air flow rate of 25.0 L/h (0.2 mol O2/h) during the whole fermentation time (232 h). As shown in figure 1, after 10 h of culture, the concentration of dissolved oxygen dropped down to zero and remained at this value the whole cultivation period. During this time the oxygen transferred to the liquid phase was totally consumed. Since this value does not give information about the rate of oxygen consumption, it was required to measure this parameter in the oulet gas of the bioreactor. This was also observed in previous reports of our group, for H. anomala, S. ludwigii and Brettanomyces intermedius yeasts when cultured under conditions similar to the ones tested [2, 3, 46]. Nevertheless, oxygen consumption by S. cerevisiae was reported to follow a different behavior after 50 h in culture. Almost the same pattern of oxygen consumption was followed by K. apiculata and H. uvarum [1, 4, 24]. The oxygen consumed by these yeasts is used in the respiration process for glucose oxidation, and also in non-respiratory pathways such as synthesis of sterols and unsaturated fatty acids, which are essential components of the cellular membrane [47].
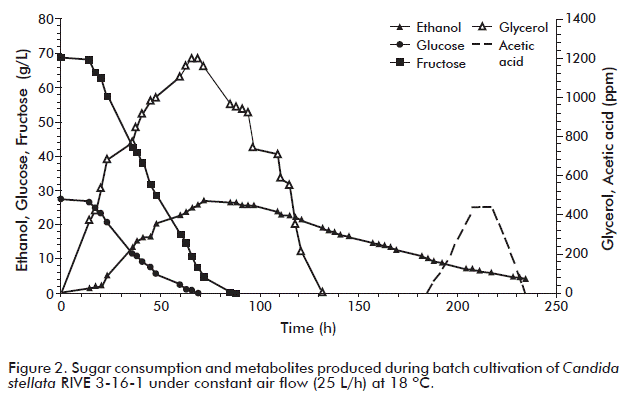
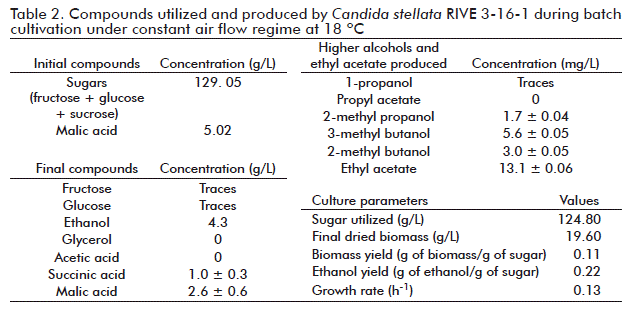
The course of the whole fermentation process in terms of sugar consumption, ethanol, glycerol and acetic acid production is shown in figure 2. Fermentation can be divided in two stages: one characterized by sugar consumption (glucose and fructose) and production of ethanol and glycerol until 70 h, followed by a second one in which fermentation byproducts (ethanol and glycerol) are consumed. Regarding yeast growth kinetics, it reached 19.6 g/L, with an specific growth rate of 0.13 h-1 at the end of culture. In this regard, Ciani et al. [12] reported that the presence of oxygen increased growth rate in approximately three-fold (from 0.05 to 0.16 h-1) when C. stellate was cultured in shake flasks at 150 rpm in synthetic medium and synthetic grape juice, as compared to anaerobic cultivation experiments. The low biomass production seen in our experiment may be due to the use of fermentable sugar by C. stellata RIVE 3-16-1 in cell maintenance reactions, as cultures were run at a low pH value (3.2-3.8). It was also observed that once sugars were consumed, ethanol and glycerol were simultaneously used as carbon source, followed by acetic acid.
As shown for sugar consumption (Figure 2), glucose was the first substrate to be depleted (up to 70 h) and then fructose (up to 90 h), this last with a higher consumption rate. This can be explained due to the fact that C. stellata is a fructophilic yeast using fructose as a preferential substrate. This sugar depletion up to 90 h was also observed previously in experiments with S. cerevisiae under similar culture conditions [24].
Ethanol and glycerol production were mainly produced in the first stage (Figure 2), the former at lower levels than in previous studies with S. cerevisiae under similar culture conditions [24]. On the other hand, acetic acid production was observed at the end of culture when ethanol was the main carbon source. In fact, it was generated by C. stellata RIVE 3-16-1 at levels three-fold lower than those attained with S. cerevisiae [24].
Simultaneously, sugar consumption resulted in maximum production of ethanol (27.0 g/L) and glycerol (1.2 g/L), these compounds serving as carbon source to maintain cell growth once sugars were depleted (Figure 2). From this stage on, the dissolved oxygen concentration dropped to zero and it remained constant toward the end of the cultivation period. Finally, acetic acid was produced toward the end of cultivation during ethanol assimilation.
In the case of glycerol, its production by yeast may constitute a response against the high osmolarity of the environment (osmoregulation) [48]. Apple juice normally contains high sugar concentrations and it can increase the osmolarity in C. stellata RIVE 3-16-1 cells, which could synthesize glycerol as compatible intracellular solute to compensate this parameter. In fermentation studies with Candida magnolia, it was shown that that cell growth and glycerol production were strongly affected by oxygen supply [49]. Apart from the effect on glycerol synthesis, the 0.13 h-1 growth rate seen (Table 2) was similar to that reported by us for other yeasts cultured under similar conditions [3, 24]. It is common knowledge that the energy generated from glucose metabolism may be mostly used in cell growth during glucose-limited and fully aerobic fermentations.
Additionally, in in-batch cultivation under constant air flow, C. stellata RIVE 3-16-1 was able to utilize 48.2 % of the total malic acid present in apple juice. This finding is interesting since yeasts with high capability to degrade malic acid are desired. The presence of oxygen in the medium would enhance its utilisation. It was also observed that degradation of malic acid started after sugar depletion and at the same time with ethanol assimilation (Figure 2 and Figure 3). This behavior was also observed in previous studies with S. cerevisiae cultured under similar conditions as used in this study [24] and, it may indicate that malic acid serves as carbon source to support the cell growth in both yeasts. Malic acid is the most important organic acid present in apple juice. It contributes to the total acidity in ciders. The decrease of its content to values lower than 5 g/L gives ciders with better sensory quality. Wine yeast in general cannot effectively degrade malic acid during alcoholic fermentation. Commercial wine yeasts (S. cerevisiae) are able to degrade only 18 % of the total malic acid [50], we previously reported degradation levels of up to 36 % of the total malic acid present in apple juice [24]. The other major metabolic compound, succinic acid, is the main organic acid generated by yeasts during metabolism of the carbon source, contributing to the taste of many alcoholic beverages [38, 44]. Here we found that C. stellata RIVE 3-16-1 produced it at 1.4 g/L. Moreover, the variability found for succinic acid synthesis throughout the entire fermentation process could be explained by its continuous excretion and reimport into yeast cells [38, 48].
Finally, other byproducts such as higher alcohols and acetate esters were also identified. They were confirmed among trace compounds remaining at the end of culture (Table 2). Probably, at the second fermentation stage (aerobic assimilation of ethanol), these higher alcohols could have served as carbon source for cell biomass increase, their low concentrations at the end of cultivation supporting this hypothesis (Table 2). From the technological point of view, fermentations should be stopped once sugars get depleted or when ethanol concentrations reach the intended value. Overall, our results are in line with common knowledge on alcoholic fermentation, regarding that excessive aeration during fermentation leads to increased cell biomass and consequently decreased ethanol production.
CONCLUSIONS
The concentration of oxygen and sugars are the most important factors that affect the metabolism of C. stellata RIVE 3-16-1. At high sugar concentration, alcoholic fermentation takes place even with insufficient oxygen amounts. In culture under agitation in Erlenmeyer flasks it was found oxygen transference is a limiting factor, probably leading to very low oxygen concentration in the medium. These oxygen-limited conditions influences the synthesis of fermentation byproducts by C. stellata RIVE 3-16-1, particularly glycerol and increases the overall production of higher alcohols (especially 3-methyl-butanol and 2-phenylethanol) and acetate esters such as ethyl acetate. Simultaneously, acetic acid diminishes under agitation, something desirable for alcoholic beverages, due to low volatile acidity. From the sensory point of view, the static culture conditions provide the best acceptability criteria, providing additional evidence for static culture as optimal for apple juice fermentation.
Conversely, constant air flow conditions supporting oxygen consumption for three days led to sugar depletion and enhanced yeast growth and fermentation rate, but affecting ethanol yields. Hence, excessive aeration should be avoided to improve ethanol yields, since C. stellata RIVE 3-16-1 normally produces ethanol, glycerol and acetic acid during aerobic fermentation, but these products are metabolized with continuous oxygen supply once the sugars are aerobically depleted. Therefore, care must be taken when main carbon sources (sugars) are exhausted, to avoid the consumption of glycerol and acetic acid as energy sources, as well as malic acid, affecting the sensory quality of the final beverage. In this case, the fermentation process should be stopped once the maximum ethanol concentration has been attained.
From the technological point of view, small amounts of oxygen would be suitable in order to control the production of ethanol and the synthesis of fermentation byproducts.
CONFLICTS OF INTEREST STATEMENT
The authors declare that there are no conflicts of interest.
REFERENCES
1. Estela-Escalante WD, Rychtera M, Melzoch K, Guerrero-Ochoa MR. Influencia de la aireación en la actividad fermentative de Kloeckera apiculata durante lafermentación de jugo de manzana. Acta Biol Colombiana. 2012;17(2):309-22.
2. Estela-Escalante WD, Rychtera M, Melzoch K, Hatta-Sakoda B, Ludeña-Cervantes Z, Sarmiento-Casavilca V, et al. Actividad fermentativa de Saccharomycodes ludwigii y evaluación de la síntesis de compuestos de importancia sensorial durante la fermentación de jugo de manzana. TIP.2011;14(1):12-23.
3. Estela-Escalante W, Rychtera M, Melzoch K, Quillama-Polo E, Hatta-Sakoda B. Estudio de la actividad fermentativa de Hansenula anomala y producción de compuestos químicos de importancia sensorial. Rev Peru Biol. 2011;18(3):325-34.
4. Estela-Escalante W, Rychtera M, Melzoch K, Hatta-Sakoda B, Quillama-Polo E, Ludeña-Cervantes Z, et al. Actividad fermentativa de Hanseniaspora uvarum y su importancia en la producción de bebidas fermentadas. Rev Soc Venezolana Microbiol. 2011;31:57-63.
5. Englezos E, Rantsiou K, Torchio F, Rolle L, Gerbi V, Cocolin L. Exploitation of the non-Saccharomyces yeast Starmerella bacillaris (synonym Candida zemplinina) in wine fermentation: Physiological and molecular characterizations. Int J Food Microbiol. 2015;199:33-40.
6. Estela-Escalante WD, Rosales-Mendoza S, Moscosa-Santillán M, González-Ramírez JE. Evaluation of the fermentative potential of Candida zemplinina yeasts for craft beer fermentation. J Inst Brew. 2016;122:530-5.
7. Estela-Escalante WD, Moscosa-Santillán M, González-Ramírez JE, Rosales-Mendoza S. Evaluation of the potential production of ethanol by Candida zemplinina yeast with regard to beer fermentation. J Am Soc Brew Chem. 2017;75(2):130-5.
8. Mora J, Mulet A. Effects of some treatments of grape juice on the population and growth of yeast species during fermentation. Am J Enol Viticult. 1991;42:133-6.
9. Jolly NP, Augustyn OPH, Pretorius IS. The occurrence of non-Saccharomyces species over three vintages in four vineyards and grape musts from four production regions of the Western Cape, South Africa. South Afr J Enol Viticult. 2003;24:35-42.
10. Ciani M, Ferraro L. Combined use of immobilized Candida stellata cells and Saccharomyces cerevisiae to improve the quality of wines. J Appl Microbiol.1998;85:247-54.
11. Magyar I, Toth T. Comparative evaluation of some oenological properties in wine strains of Candida stellata, Candida zemplinina, Saccharomyces uvarum and Saccharomyces cerevisiae. Food Microbiol. 2011;28:94-100.
12. Ciani M, Ferraro L, Fatichenti F. Influenceof glycerol production on the aerobic and anaerobic growth of the wine yeast Candida stellata. Enz Microb Technol. 2000;27:698-703.
13. Rodicio R, Heinisch JJ. Sugar metabolism by Saccharomyces and non-Saccharomyces yeasts. In: Helmut-Konig H, Unden G, Frohlich J (Editors). Biology of microorganisms on grapes, in must and inwine. Berlin Heidelberg: Springer-Verlag;2009. p. 113-34.
14. Van Dijken JP, Scheffers WA. Redox balances in the metabolism of sugars by yeasts. FEMS Microbiol Rev. 1986;32:199-224.
15. Verduyn C. Physiology of yeasts inrelation to biomass yields. Antonie van Leeuwenhoek. 1991;60:325-53.
16. Ribereau-Gayon P, Dubourdieu D, Doneche B, Lonvaud A. Biochemistry of alcoholic fermentation and metabolic pathways of wine yeasts. In: Handbook of Enology. The Microbiology of Wine and Vinifications. New York: John Wiley and Sons; 2000. p. 51-74.
17. Suomalainen H. Yeast esterase and aroma esters in alcoholic beverages. J Inst Brew. 1981;87:296-300.
18. Nordstrom K. Formation of ethyl acetate in fermentation with brewer’s yeast. III. Participation of coenzyme A. J Inst Brew. 1962;68:398-407.
19. Yoshioka K, Hashimoto H. Cellular fatty acid and ester formation by brewers’ yeast. Agr Biol Chem. 1983;47:2287-94.
20. Hammond JRM. Brewer’s yeasts. In: Rose AH, Harrison JS (Editors). The Yeast.Yeast technology (vol. 5). London: Academic Press; 1993. p. 7-67.
21. Barker RL, Irwin JA, Murray CR. The relationship bewteen fermentation variables and flavour volatiles by direct gas chromatographic injection of beer. Master Brewers Assoc Am Tech Quart.1992;29:11-7.
22. Downing DL. Apple Cider. In: DonaldLD (Editor). Processed Apple Products. NewYork: Van Nostrand Reinhold Publisher. USA; 1989. p. 169-188.
23. Su SK, Wiley RC. Changes in applejuice flavor compounds during processing. J Food Sci. 1998;63(4):688-69.
24. Estela-Escalante WD, Rychtera M, Melzoch K, Hatta-Sakoda B. Effect of aeration on the fermentative activity of Saccharomyces cerevisiae cultured in apple juice. Rev Mexicana Ing Quím. 2012;11(2):211-26.
25. Ortega C, Lopez R, Cacho J, Ferreira V. Fast analysis of important wine volatile compounds: Development and validation of a new method based on gas chromatography-flame ionization detection analysis of dichloromethane microextracts. J Chrom A. 2001;923:205-14.
26. Meilgaard M, Civille GV, Carr BT. Sensory Evaluation Techniques. 4rd edition. Boca Raton, New York: CRC Press;2006. p. 22-35.
27. Gupta A, Rao GA. Study of oxygen transfer in shake flasks using a noninvasive oxygen sensor. Biotechnol Bioeng. 2003;84(3):351-8.
28. Cabranes C, Moreno J, Mangas JJ. Cider production with immobilized Leuconostoc oenos. J Inst Brew. 1998;4:127-30.
29. Souci SW, Fachmann W, Kraut H. Juices from fruits and berries: Food composition and nutrition tables. 6th edition. Stuttgart: Scientific publishers; 2000. p. 13-52.
30. Wang L, Xu Y, Zhao G, Li J. Rapid analysis of flavor volatiles inapple wine using Headspace Solid-Phase microextraction. J Inst Brew. 2004;110(1):57-65.
31. Picinelli A, Suarez B, Moreno J, Rodríguez R, Caso-García LM, Mangas JJ. Chemical characterization of Asturian cider. J Agr Food Chem. 2000;48:3997-4002.
32. Suarez-Valles B, Pando-Bedrinana R, Fernandez-Tascon N, Gonzales-Garcia A, Rodríguez-Madrera R. Analytical differentiation of cider inoculated with yeast (Saccharomyces cerevisiae) isolated from Asturian (Spain) apple juice. LWT-Food Sci Technol. 2005;38:455-61.
33. Jarvis B, Forster MJ, Kinsella WP. Factors affecting the development of cider flavour. J Appl Bacteriol. 1995;79:5S-18S.
34. Ciani M, Maccarelli F. Oenological properties of non-Saccharomyces yeasts associated with winemaking. World J Microbiol Biotechnol. 1998;14:199-203.
35. Prior BA, Toh TH, Jolly N, Baccari C, Mortimer RK. Impact of yeast breeding for elevated glycerol production on fermentation activity and metabolite formation in Chardonnay. South Afr J Enology Viticulture. 2000;21:92-9.
36. Nieuwoudt HH, Prior BA, Pretorius IS, Bauer FF. Glycerol in South African table wines: an assessment of its relationship to wine quality. South Afr J Enol Viticult. 2002;23:22-30.
37. Swiegers JH, Pretorius IS. Yeast modulation of wine flavour. Adv Appl Microbiol. 2005;57:131-75.
38. Swiegers JH, Bartowsky EJ, HenschkePA, Pretorius IS. Yeast and bacterial modulation of wine aroma and flavour. Austr J Grape Wine Res. 2005;11:139-73.
39. Rapp A, Mandery H. New progress in wine and wine research. Experientia. 1986;42(8):873-84.
40. Ribereau-Gayon P, Dubourdieu D, Doneche B. Biochemistry of alcoholic fermentation and metabolic pathways of wine yeasts. In: Handbook of enology. The microbiology of wine and vinifications. Chichester: John Wiley and Sons, Ltd;2006. p. 74-7.
41. Ribereau-Gayon P. Wine aroma. In: Charalambous G, Inglett GE (Editors). Flavour of foods and beverages. NewYork: Academic Press; 1978. p. 362-71.
42. Dequin S, Salmon JM, Nguyen HV. Wine yeasts. In: Boekhout T, Robert V(Editors). Yeasts in food-beneficial and detrimental aspects. Hamburg: Behr’s Verlag; 2003. p. 389-412.
43. Dubois P. Les aromes des vins et leurs defauts. Revue Francaise d’Oenologie.1994;145:27-40.
44. Coulter AD, Godden PW, Pretorius IS. Succinic acid-How it is formed, what is its effect on titratable acidity, and what factor sinfluence its concentration in wine? Austr New Zealand Wine Ind J. 2004;19:16-25.
45. Furukawa K, Heinzle E, Dunn IJ. Influence of oxygen on the growth of Saccharomyces cerevisiae in continuous culture. Biotechnol Bioeng. 1983;25(10):2293-317.
46. Estela-Escalante WD, Rychtera M, Melzoch K, Torres-Ibáñez F, Calixto-Cotos R, Bravo-Araníbar N, et al. Efecto de la aireación en la producción de compuestos volátiles por cultivo mixto de Brettanomyces intermedius y Saccharomyces cerevisiae durante la fermentación de sidra. TIP.2014;17(1):5-14.
47. Rosenfeld E, Beauvoit B. Role of the non-respiratory pathways in the utilization of molecular oxygen by Saccharomyces cerevisiae. Yeast. 2003;20:1115-44.
48. Hohmann S. Osmotic stress signaling and osmoadaptation in yeasts. Microbiol Mol Biol Rev. 2002;66:300-72.
49. Sahoo DK, Agarwal GP. Effect of oxygen transfer on glycerol biosynthesis by an osmophilic yeast Candida magnoliae I2B. Biotechnol Bioeng. 2002;78(5):545-55.
50. Redzepovic S, Orlic S, Majdak A, Kozina B, Volschenk H, Viljoen-Bloom M. Differential malic acid degradation by selected strains of Saccharomyces during alcoholic fermentation. Int J Food Microbiol. 2003;83:49-61.
Received in October, 2017.
Accepted in March, 2018.
Waldir D Estela-Escalante. Escuela Profesional de Ingeniería Agroindustrial. Facultad de Química e Ingeniería Química, Universidad Nacional Mayor de San Marcos. Av. Universitaria s/n. Lima 1, Lima, Perú. E-mail: waldir.estela@unmsm.edu.pe.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}