










My SciELO
 Custom services
Custom servicesServices on Demand
Article
 Portuguese (pdf)
Portuguese (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista Cubana de Plantas Medicinales
On-line version ISSN 1028-4796
Rev Cubana Plant Med vol.22 no.1 Ciudad de la Habana Jan.-Mar. 2017
ARTÍCULO ORIGINAL
Caracterização e avaliação morfológica da parte aérea de acessos de Psychotria ipecacuanha (Brot.) Stokes (IPECA)
Caracterización y evaluación morfológica de partes aéreas de accesiones de Psychotria ipecacuanha (Brot.) Stokes (IPECA)
Characterization and morphological evaluation of aerial parts of accessions of Psychotria ipecacuanha (Brot.) Stokes (IPECA)
Raphael Lobato Prado Neves,I Osmar Alves Lameira,II Maria do Socorro Padilha de Oliveira,II Fernanda Ilkiu Borges de Souza,II Lucila Elizabeth Fragoso Monfort,I Ruy Guilherme CorreiaI
IUniversidade Federal Rural da Amazônia-UFRA. Brasil.
IIEmpresa brasileira de pesquisa agropecuária-EMBRAPA. Brasil.
RESUMO
Introdução: a espécie Psychotria ipecacuanha (Brotero) Stokes, Rubiaceae, conhecida popularmente como ipeca, ipecacuanha ou poaia, encontra-se ameaçada de erosão genética e em vias de extinção.
Objetivos: caracterizar e avaliar acessos do Banco Ativo de Germoplasma de Psychotria ipecacuanha por caracteres morfológicos, e identificar descritores.
Métodos: foram estudados 17 acessos de Psychotria ipecacuanha, conservados in vivo na Embrapa Amazônia Oriental para 19 caracteres morfológicos da parte aérea da planta, sendo treze qualitativos e seis quantitativos. Desse total doze foram submetidos às análises multivariadas. A seleção dos descritores foi realizada pela análise de componentes principais.
Resultados: os acessos de Psychotria ipecacuanha apresentaram variações para boa parte dos caracteres, sendo sete qualitativos não variáveis. Dois caracteres foram considerados redundantes e dez selecionados como descritores. As dissimilaridades genéticas variaram de 0,19 a 0,77 com média de 0,55 e permitiram separar os acessos em dois com vários subgrupos e seis grupos divergentes pelos métodos UPGMA e de Tocher, respectivamente. Os caracteres "Altura da planta" e "Altura da primeira ramificação" foram os que mais contribuíram para a divergência entre os acessos.
Conclusões: os acessos de Psychotria ipecacuanha possuem ampla variação morfológica para a parte aérea da planta com dez caracteres sendo considerados descritores para essa espécie. Estas informações são inéditas para a espécie e servirão de base para o manejo do banco de germoplasma, na seleção de indivíduos de interesse dos programas de melhoramento e para fornecer informações fundamentais que facilitarão a identificação de espécimes em ecossistemas nativos ou plantados.
Palavras chave: componentes principais; variação morfológica; agrupamentos; plantas medicinais; Rubiaceae.
RESUMEN
Introducción: la especie Psychotria ipecacuana (Brotero) Stokes, Rubiaceae, popularmente conocida como ipeca, ipecacuanha o poaia, se ve amenazada por la erosión genética y en peligro de extinción.
Objetivos: identificar y evaluar accesos del Banco Activo de Germoplasma de Psychotria ipecacuana por caracteres morfológicos, e identificar los descriptores.
Métodos: se estudiaron 17 accesiones de Psychotria ipecacuana, conservado in vivo en Embrapa Amazonia Oriental, para 19 caracteres morfológicos de la parte aérea de la planta, siendo trece cualitativo y seis cuantitativo. De este total doce fueron sometidos a análisis multivariante. La selección de descriptores se llevó a cabo mediante el análisis de componentes principales.
Resultados: los accesos de Psychotria ipecacuana fluctuaron para la mayoría de los caracteres, sendo siete variables cualitativas y no variables. Dos caracteres se consideraron redundante y diez seleccionados como descriptores. La disimilitud genética varió desde 0,19 hasta 0,77 con una media de 0,55 y se dejó separar en dos grupos con diversos subgrupos y seis grupos diferentes por métodos UPGMA y Tocher, respectivamente. Los caracteres "altura de las plantas" y "altura de la primera rama" fueron los principales contribuyentes a la discrepancia entre los accesos.
Conclusiones: los accesos de Psychotria ipecacuana tienen una amplia variación morfológica de la parte aérea de la planta con diez caracteres fueran considerados descriptores para esta especie. Estos datos son inéditos para la especie y forman la base para la gestión de banco de germoplasma, en la selección de caracteres de interés para los programas de mejoramiento y para proporcionar información clave que facilitará la identificación de especímenes en los ecosistemas naturales o plantados.
Palabras clave: componentes principales; variación morfológica; grupos; plantas medicinales; Rubiaceae.
ABSTRACT
Introduction: The species Psychotria ipecacuana (Brotero) Stokes, Rubiaceae, commonly known as ipeca, ipecacuanha or poaia, is under threat of genetic erosion and in danger of extinction.
Objectives: Identify and evaluate the morphological characters of accessions of Psychotria ipecacuana and identify their descriptors.
Methods: A study was conducted of 17 accessions of Psychotria ipecacuana preserved in vivo at EMBRAPA Eastern Amazonia for 19 morphological characters of the aerial parts of the plant, thirteen qualitative and six quantitative. Of this total, twelve were subjected to multivariant analysis. Principal component analysis was used to select the descriptors.
Results: Accessions of Psychotria ipecacuana fluctuated for most characters, and seven were qualitative and nonvariable. Two characters were considered redundant and ten were selected as descriptors. Genetic dissimilarity ranged from 0.19 to 0.77 with a mean value of 0.55, and could be separated into two groups with various subgroups and six different groups by the UPGMA and Tocher methods, respectively. The characters "plant height" and "first branch height" were the main contributors to the discrepancy between the accessions.
Conclusions: Accessions of Psychotria ipecacuana display great morphological variation in the aerial part of the plant, with ten characters selected as descriptors for the species. These data about the species had never been published before, and they constitute the basis for management at the germplasm bank when selecting characters of interest for improvement programs as well as to provide key information facilitating the identification of specimens from natural or planted ecosystems.
INTRODUÇÃO
A espécie Psychotria ipecacuanha (Brot.) Stokes, Rubiaceae, conhecida popularmente como ipeca, ipecacuanha ou poaia, encontra-se ameaçada de erosão genética e em vias de extinção.1 Os fatores que têm contribuído para essa ameaça envolvem a exploração intensa a partir do século XVI, devido elevado valor comercial de suas raízes; desmatamento da Mata Atlântica e a abertura de novas fronteiras agrícolas diminuindo suas áreas de ocorrência natural; a forma extrativista com plantios de P. ipecacuanha inexpressivos e sem o manejo adequado; e a dificuldade do cultivo convencional da espécie, frente à grande demanda de mercado.2,3 Além disso, a espécie apresenta escassez de estudos que possam orientar plantios racionais, principalmente aqueles que envolvam a geração de conhecimentos sobre o germoplasma conservado.
Há registros de que nas raízes de P. ipecacuanha hasão encontrados valiosos alcaloides como, a psicotrina, emetina e cefalina, que conferem à espécie poderes eméticos, amebicida, expectorante e anti-inflamatório.2 Essas propriedades medicinais são conhecidas e utilizadas pelas populações nativas, desde antes do descobrimento do Brasil, os quais foram repassados aos colonizadores europeus, contribuindo para a espécie integrar a pauta de produtos tropicais exportados pelo Brasil no período colonial, sendo considerada como a espécie de maior valor medicinal da época.4-7
Com vista à avançar nos estudos de conservação da espécie, em 1989, foi estabelecido o Banco Ativo de Germoplasma (BAG) de P. ipecacuanha, na Embrapa Amazônia Oriental, Belém-PA, sendo este o único banco ativo de P. ipecacuanha existente. Porém, nesse banco os acessos ainda não foram caracterizados e avaliados morfologicamente.
A caracterização e a avaliação morfológica se mostram vantajosas por manter as informações dos BAG's atualizadas e disponíveis para os mais diversos usos, além de apresentar baixo custo em comparação com as demais técnicas, pelo fato de ser elaborada in vivo e em curto tempo, não precisando de equipamentos complexos ou análises laboratoriais e que elevam o dispêndio financeiro. Essas atividades auxiliam na seleção prévia de indivíduos de interesse, onde descritores bem definidos podem garantir características hereditárias próprias e fornecer os elementos para manutenção da pureza vegetal, além de haver facilidade em ser replicada periodicamente.8
Visando contribuir com informações que servirão de base para a seleção de indivíduos de interesse dos programas de melhoramento genético; além de fornecer informações fundamentais que facilitarão a identificação de indivíduos em ecossistemas nativos ou plantados; este estudo inédito para a espécie em foco, teve o objetivo de caracterizar e avaliar acessos pertencentes ao banco ativo de germoplasma de P. ipecacuanha, e identificar descritores morfológicos para a espécie.
MÉTODOS
Área do estudoO estudo foi realizado no Banco Ativo de Germoplasma de P. ipecacuanha, situado no Horto de Plantas Medicinais da Embrapa Amazônia Oriental, com altitude de 10 m e temperatura média anual de 30 °C. O clima em Belém, segundo a classificação de Köppen (1900-1936) é do tipo Afi, ou seja, clima tropical chuvoso de monção. A pluviosidade média anual é de 2.858,7 ± 76,6 mm/ano com maior volume no período chuvoso (dezembro a maio), correspondendo a 71,2 % do total anual, enquanto que os 28,8 % restantes são distribuídos nos meses de junho a novembro.9
Germoplasma avaliado
Foram avaliados 17 acessos, conservados in vivo no banco ativo de germoplasma de Psychotria ipecacuanha (Brot.) Stokes. P. ipecacuanha pertence à família Rubiaceae, registrado no herbário IAN, Belém, Pará (4093 IAN); em cada acesso foram selecionados cinco indivíduos para a caracterização morfológica. Os acessos se encontram em ambiente onde há o controle de pragas e doenças, adubação orgânica e intensidade luminosa com sombrite 50 %.
Os acessos foram coletados em diferentes localidades de três Estados do Brasil, sendo seis em Mato Grosso (590, 595, 596, 602, 607 e 707); um acesso em Minas Gerais (753); e dez acessos em Rondônia (610, 612, 819, 826, 828, 829, 833, 834, 836 e 837).
Caracterização morfológica
Os caracteres morfológicos qualitativos utilizados foram: Forma foliar (FF); Base foliar (BF); Ápice foliar (AF); Cor da folha apical (CDFA); Cor do pecíolo (CDP); Cor da nervura foliar (CDN); Tipo de caule (TC); Tipo de folha (TF); Filotaxia (F); Consistência da folha (CDF); Tipo de venação foliar (TDVF); Tipo de estípula (TDE); Posição do pecíolo (PDP). Os caracteres forma foliar, base foliar e ápice foliar foram aplicados com base em Carlton;10 enquanto as cores da folha apical, do pecíolo e da nervura foliar foram observadas com o auxílio da carta de cores Standard Leaf Color Charts.11
Seleção de descritores e divergência genética entre os acessos
Os caracteres morfológicos quantitativos avaliados foram: Altura da planta (AP), em centímetros; Altura da primeira ramificação (APR), em centímetros; Número de nós na haste principal (NNHP), pela contagem dos nós existentes na planta; Largura da folha (LF); Comprimento da folha (CF); e Comprimento do pecíolo (CP), em centímetros. Com exceção do número de nós na haste principal, os demais foram aferidos com auxílio de régua e paquímetro.
Após tabulação dos dados, foram eliminados os caracteres qualitativos redundantes ou invariáveis e os demais foram utilizados juntos com os quantitativos para a seleção dos descritores com base em dois procedimentos com o auxílio do software Genes:12 o primeiro procedimento foi o de seleção direta, proposto por Jolliffe,13,14 neste caso foram eliminados todos os caracteres que apresentaram maior coeficiente de ponderação em valor absoluto (autovetor) no componente principal de menor autovalor, partindo do último componente até aquele cujo autovalor não excedesse a 0,70. O segundo foi o a seleção com reanálise, sugerida por Cury,15 em que a cada caráter descartado, foi realizada nova análise com os caracteres remanescentes, examinando os coeficientes de correlação do caráter sugerido para descarte com os demais caracteres, sendo finalizada quando o caráter a ser descartado se mostrou altamente correlacionado com pelo menos um caráter já descartado. O descarte final foi realizado com base na informação obtida simultaneamente nos dois procedimentos.
Os dez descritores selecionados foram utilizados, no mesmo software, para a obtenção das dissimilaridades entre cada par de acesso com base na distância euclidiana média padronizada, uma vez que os acessos encontram-se estabelecidos sem obedecer a nenhum delineamento experimental, assim como os caracteres em diferentes medidas e escalas. Esta análise foi executada com base na média de cada caráter.
A matriz de distâncias obtidas foi utilizada na formação dos agrupamentos pelos métodos Unweighted Paired Group Method Using Arithmetic Averages (UPGMA),16 de otimização de Tocher e pela análise de componentes principais.
RESULTADOS
Caracterização morfológica
Dos treze caracteres qualitativos foram eliminados sete caracteres por não terem apresentado variação entre os acessos: Tipo de caule (Ereto); Tipo de folha (Simples); Filotaxia (Opostas decussadas); Consistência da folha (Membranosa); Tipo de venação foliar (Peninérvea); Tipo de estípula (Interpeciolada); Posição do pecíolo (Oposto), enquanto os demais mostraram duas variações (tabela 1).
Nos acessos, o caráter Forma foliar (FF) variou de "Obovada" e "Elíptica", sendo 10 acessos (590, 596, 612, 707, 753, 819, 828, 834, 836 e 837) de forma foliar obovada e sete (595, 602, 607, 610, 826, 829 e 833) elíptica. O caráter Base foliar (BF) obteve duas variações "Revoluta" e "Cuneada", onde 13 acessos (595, 602, 607, 610, 612, 819, 826, 828, 829, 833, 834, 836 e 837) foram classificados como "Cuneada" e quatro (590, 596, 707 e 753) como "Revoluta". Para o caráter Ápice foliar (AF), a variação foi de "Acuminado" e "Agudo", sendo que 13 acessos (590, 595, 596, 607, 610, 612, 707, 753, 819, 826, 828, 829 e 833) foram classificados como "Agudo" e quatro (602, 834, 836 e 837) como "Acuminado".
Os caracteres Cor da folha apical (CDFA), Cor do pecíolo (CDP) e Cor da nervura foliar (CDN) apresentaram duas tonalidades "Verde escuro" e "Verde claro", mas de intensidades distintas (tabela 1). Para a Cor da folha apical 10 acessos (607, 612, 753, 819, 826, 829, 833, 834, 836 e 837) apresentaram folha apical verde escuro (7.5 GY 4/4) e sete acessos (590, 595, 596, 602, 610, 707 e 828) verde claro (7.5GY 4.5/5). No caráter Cor do pecíolo, oito acessos (590, 595, 602, 607, 612, 826, 828 e 829) foram classificados como "verde escuro" (7.5GY 3.5/4) e nove acessos (596, 610, 707, 753, 819, 833, 834, 836 e 837) como "verde claro" (7.5GY 4.5/4). No caso da Cor da nervura foliar cinco acessos (590, 595, 596, 602 e 834) foram classificados como "verde escuro" (7.5GY 5/7) e 12 acessos (607, 610, 612, 707, 753, 819, 826, 828, 829, 833, 836 e 837) como "verde claro" (7.5GY 6/7).
Seleção dos descritores e divergência entre os acessos
Dos doze caracteres avaliados foram sugeridos para descarte dois deles: um qualitativo, o Ápice foliar (AF), e um quantitativo, o Comprimento da folha (CF), os quais se mostraram redundantes nos dois procedimentos utilizados. Os demais caracteres foram sugeridos como descritores para a espécie.
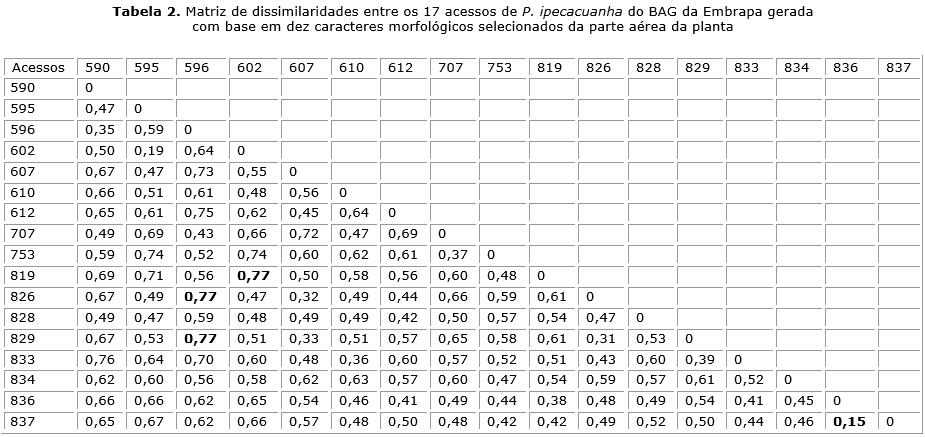
As dissimilaridades obtidas entre os 136 pares de acessos variaram de 0,15 a 0,77 (tabela 2) com média de 0,55. Os acessos mais semelhantes, ou seja, de maior similaridade foram os acessos 836 × 837, que apresentaram a menor distância (0,15), seguido pelos acessos 595 × 602, com 0,19 de dissimilaridade. Os pares de acessos mais divergentes, ou seja, de maior dissimilaridade foram 602 × 819, 596 × 826 e 596 × 829, todos com a maior distância (0,77).
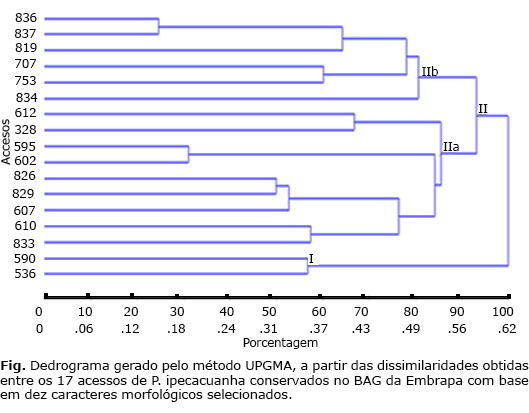
Com base no método UPGMA os 17 acessos formaram dois grupos divergentes: o I contendo os acessos 590 e 596, coletados em diferentes Municípios do Estado de Mato Grosso e o II pelos demais acessos (fig.). No grupo II os acessos foram organizados em vários subgrupos, com dois subgrupos apresentando consistência acima de 90 % (IIa e IIb). No subgrupo IIa ficaram nove acessos (612, 828, 595, 602, 826, 829, 607, 610 e 833) a maioria coletado no Município de Costa Marques, Ro. Enquanto no subgruo IIb foram agrupados seis acessos de diferentes localidades (836, 837, 819, 707, 753 e 834).
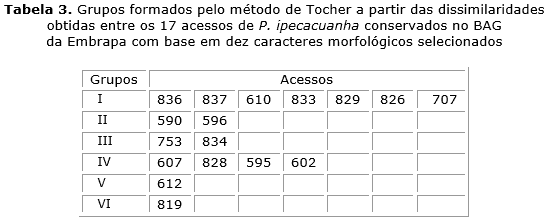
Pelo método de Tocher houve a formação de seis grupos divergentes entre os 17 acessos (tabela 3). No grupo I foram agrupados sete acessos (836, 837, 610, 833, 829, 826 e 707) de diferentes localidades; o grupo II contendo dois acessos (590 e 596), ambos coletados em Mato Grosso, sendo esse coincidente ao grupo I formado pelo método UPGMA; o grupo III formado também por dois acessos (753 e 834) coletados em diferentes Estados; o grupo IV formado por quatro acessos (607, 828, 595 e 602), três deles coletados em Mato Grosso; e os grupo V e VI constituído por um acesso, 612, de Costa Marques e 819 de Cacoal, respectivamente.
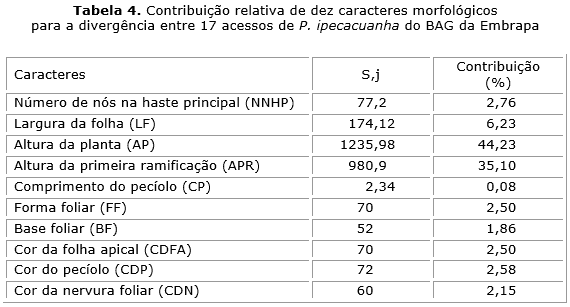
A Altura da planta (AP) e Altura da primeira ramificação (APR) foram os caracteres que mais contribuíram para a divergência entre os 17 acessos com mais de 44 % e 35 %, respectivamente (tabela 4). Por outro lado, a menor contribuição ocorreu no caráter comprimento do pecíolo (CP) com 0,08 %.
DISCUSSÃO
Caracterização morfológica
Assis & Giulieti17 em um estudo comparativo de populações de P. ipecacua, afirmaram que as variações associadas ao comprimento e largura das folhas, estão expressas na variação das folhas, na qual variaram desde elípticas até ovais, oblongo-lanceoladas ou obovais. Observaram também que não houve correlação entre os padrões de morfologia foliar encontrados e a distribuição geográfica.
No presente estudo as variações morfológicas da forma da folha estão entre Obovada e Elíptica, sendo que a maioria (dez acessos) foi representada por Obovada e os demais, como elíptica. Este resultado está de acordo com a caracterização morfológica feita por Assis & Giulieti.17
Seleção de descritores e divergência entre os acessos
Para a seleção de descritores, todo caráter deve apresentar uma parcela de contribuição na variação do germoplasma analisado. Mas há uma tendência de que o aumento do número de descritores avaliados ocasione a presença de caracteres redundantes, posto que esses caracteres quase sempre estão associados a outros.16 A eliminação dos redundantes é uma decisão vantajosa, pois reduz o trabalho sem ocasionar perda na precisão da caracterização, especialmente se esses caracteres forem de difícil mensuração e apresentarem baixa variabilidade,18 por isso, neste trabalho foram eliminados todos os caracteres redundantes.
A análise de componentes principais vem se destacando como a metodologia mais empregada em bancos e ou coleções de germoplasma, pois além de identificar os caracteres mais importantes na contribuição de variação total disponível entre os indivíduos analisados, fornece indicação para eliminar os que pouco contribuem.19
Assis & Giulietti17 realizando avaliação de 97 populações de P. ipecacuanha de diversas procedências, baseadas no exame de exsicatas provenientes de diversos herbários nacionais e estrangeiros, afirma que o primeiro componente principal foi a simetria da folha, que incluiu o comprimento e a largura das folhas e que representou 72,4 % do total. O segundo componente principal foi a distância entre os entrenós e que representou 13,1 % da variabilidade total.
A diversidade genética pode estar associada à distância geográfica. Neste estudo, os acessos com maior dissimilaridade (602 × 819, 596 × 826 e 596 × 829) apresentam procedência de diferentes estados, sendo 602 e 596 procedentes do estado do Mato Grosso e os acessos 819, 826 e 829 do estado de Rondônia. O mesmo ocorreu para os acessos que apresentaram a menor dissimilaridade, ou seja, os mais semelhantes, sendo os acessos 836 × 837 procedentes de Rondônia e 595 × 602 do Mato Grosso. Fatores, como a seleção, deriva genética, fluxo gênico, entre outros, podem atuar de forma conjunta ou isolada, em diferentes intensidades e determinar a existência ou não de algum paralelismo entre essas medidas.20
No subgrupo II bonde foram agrupados seis acessos, constata-se que quatro deles (836, 837, 819 e 834) são provenientes do estado de Rondônia, o 707 proveniente do Mato Grosso e o 753 proveniente de Minas Gerais. Já no subgrupo IIa formado pornove acessos, verifica-se que seis acessos (612, 828, 826, 829, 610 e 833) são procedentes de Rondôniae três acessos (595, 602 e 607) do Mato Grosso. Pode-se perceber que apesar de diferentes procedências, houve o agrupamento destes acessos. O aumento da distância geográfica nem sempre implica em maior divergência genética, pois a deriva genética e a seleção em diferentes ambientes podem contribuir muito mais para a divergência do que a separação geográfica.21
Pelo método de Tocher observou-se alguma semelhança na formação do grupos com as demais análises, ressalta-se a formação de seis grupos, sendo o grupo I com cinco acessos (836, 837, 610, 833 e 826) provenientes de Rondônia e apenas o 707 proveniente do Mato Grosso. O grupo II com os acessos 590 e 596, ambos do estado do Mato Grosso, semelhante ao encontrado no grupo 2 da Distância Euclidiana Média. O grupo III ficou agrupado por dois acessos, 753 (estado de Minas Gerais) e 834 (estado de Rondônia). O grupo IV pelos acessos 607, 595 e 802 do Mato Grosso e 828 de Rondônia. E aqueles com maior dissimilaridade formando grupos distintos, o grupo V formado pelo acesso 612 (proveniente do Mato Grosso) e o grupo VI pelo acesso 819 (proveniente de Rondônia).
Neste trabalho, os descritores que mais contribuíram para a divergência entre os acessos foram Altura da planta (AP) e Altura da primeira ramificação (APR), ressalta-se que as procedências deste presente estudo abordaram três estados brasileiros e com espécimes in vivo, e no trabalho de Assis & Giulietti17 foram avaliadas procedências de outros países através de exsicatas.
Este estudo caracterizou e avaliou a variabilidade morfológica dos acessos de P. ipecacuanha, identificou e agrupou os descritores com menor e maior contribuição. Estas informações são inéditas para a espécie e servirão de base para a seleção de indivíduos de interesse dos programas de melhoramento, além de fornecer informações fundamentais que facilitarão a identificação de espécimes em ecossistemas nativos ou plantados.
REFERÊNCIAS
2. Lameira OA. Cultivo da Ipecacuanha Psychotria ipecacuanha (Brot.) Stokes. Belém: Embrapa Amazônia Oriental. 2002.
3. Rocha TT, Lameira OL. Avaliação do período de floração e frutificação do BAG ipecacuanha. In: Anais do 15° Seminário de Iniciação Científica da Embrapa. Belém: Embrapa Amazônia Oriental. 2011.
4. Miranda GO. A poaia: um estudo em Barra do Bugres [monografia]. Cuiabá: Universidade Federal de Mato Grosso. 1983.
5. Akerele O, Heywood V, Synge H. The conservation of medicinal plants. Cambridge: Cambridge University Press. 1991.
6. Assis MC. Aspectos taxonômicos, anatômicos e econômicos da "ipeca" Psychotria ipecacuanha (Brot.) Stokes. (Rubiaceae) [dissertação]. São Paulo: Universidade de São Paulo. 1992.
7. Corrêa MX. Ensaio bibliográfico sobre a Economia da Poaia na Zona da Mata Mineira. In: Anais do XX ciclo de estudos históricos. Ilhéus: Universidade Estadual de Santa Cruz. 2010.
8. Silva ML. Cultivo agroflorestal de Psychotria ipecacuanha (Brot.) Stokes no território do baixo sul da bahia [dissertação]. Feira de Santana: Universidade Estadual de Feira de Santana. 2014.
9. Silva Júnior JA, Costa ACL, Pezzuti JCB, Costa RF, Galbraith D. Análise da Distribuição Espacial do Conforto Térmico na Cidade de Belém, PA no Período Menos Chuvoso. Revista Brasileira de Geografia Física. 2012;2:218-32.
10. Carlton WM. Laboratory Studies in General Botany. New York: Ronald Press Company. 1961.
11. Kiuchi T, Yazawa H. Standard Leaf Color Charts. Tokyo: National Institute of Agricultural Sciences. 1972.
12. Cruz CD. Programa Genes: Biometria. Viçosa: Editora UFV. 2006.
13. Jolliffe IT. Discarding variables in a principal component analysis. I: artificial data. Journal of the Royal Statistical Society Series C. 1972;21(2):160-73.
14. Jolliffe IT. Discarding variables in a principal component analysis. II: real data. Journal of the Royal Statistical Society Series C. 1973;22(1):21-31.
15. Cury R. Dinâmica evolutiva e caracterização de germoplasma de mandioca (Manihot esculenta Crantz) na agricultura autóctone do Sul do Estado de São Paulo [dissertação]. Piracicaba: Escola Superior de Agricultura Luiz de Queiroz, Universidade de São Paulo. 1993.
16. Cruz CD, Regazzi AJ, Carneiro PCS. Modelos Biométricos Aplicados ao Melhoramento Genético. Viçosa: editora UFV. 2004.
17. Assis MC, Giulietti AN. Diferenciação morfológica e anatômica em populações de "ipecacuanha" - Psychotria ipecacuanha (Brot.) Stokes (Rubiaceae). Revista Brasileira de Botânica. 1999;22(2):205-16.
18. Daher RF. Diversidade morfológica e isoenzimática em capim elefante (Pennisetum purpureum Schum.) [dissertação]. Viçosa: Universidade Federal de Viçosa. 1993.
19. Pereira VA. Utilização de análise multivariada na caracterização de germoplasma de mandioca ( Manihot esculenta Crantz.) [tese]. Piracicaba: Escola Superior de Agricultura Luiz de Queiroz, Universidade de São Paulo. 1989.
20. Alves RM. Caracterização genética de populações de cupuaçuzeiro, Theobroma grandiflorum (Will ex Spreng) Schum., por marcadores microssatélites e descritores botânico-agronômicos [tese]. Piracicaba: Escola Superior de Agricultura Luiz de Queiroz, Universidade de São Paulo. 2002.
21. Dias LAS. Divergência genética e fenética multivariada na predição de híbridos e preservação de germoplasma de cacau (Theobroma cacao L.) [tese]. Piracicaba: Escola Superior de Agricultura Luiz de Queiroz, Universidade de São Paulo. 1994.
Recibido: 15 de febrero de 2016.
Aprobado: 27 de enero de 2017.
Raphael Lobato Prado Neves. Universidade Federal Rural da Amazônia. Correo electrónico: raphael.lobato@outlook.com


{kind=link}
{kind=link}