










My SciELO
 Custom services
Custom servicesServices on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkCuban Journal of Agricultural Science
On-line version ISSN 2079-3480
Cuban J. Agric. Sci. vol.49 no.1 Mayabeque Jan.-Mar. 2015
ORIGINAL ARTICLE
Nutritive value improvement of seasonal legumes Vigna unguiculata, Canavalia ensiformis, Stizolobium niveum, Lablab purpureus, through processing their grains with Trichoderma viride M5-2
Mejora del valor nutritivo de las leguminosas temporales Vigna unguiculata, Canavalia ensiformis, Stizolobium niveum, Lablab purpureus mediante el procesamiento de sus granos con Trichoderma viride M5-2
Elaine Valiño,I Lourdes Savón,I A. Elías,I Marleny Rodriguez,II Nereyda Albelo,I
IInstituto de Ciencia Animal, Apartado Postal 24, San José de las Lajas, Mayabeque, Cuba.
IIInstituto de Materiales y Reactivos para la electrónica (IMRE), Universidad de La Habana, Cuba.
ABSTRACT
In order to improve the nutritive value of seasonal legumes through processing their grains with Trichoderma viride M5-2, several experiments were conducted at laboratory scale for producing inocula of this mutant cell, without adding nitrogen and minerals sources for different fermentation variants. Completely randomized designs with 4x6 and 4x2 factorial arrangement and three repetitions were used. Factors varied depending on the variant analysis. Factors were fermentation substrates (Vigna unguiculata, Canavalia ensiformis, Stizolobium niveum, and Lablab purpureus) and fermentation times (0, 24, 48, 72, 96 and 120 h). The experiments were conducted in erlenmeyer flasks of 500 mL with 10g of samples at 70% humidity. Samplings were conducted every 24 h to determine cellulolitic activity (endo â1-4 glucanase and exo â1-4 glucanase), mineral composition (Ca, Mg, P, K, ash), and bromatological composition (CP, TP, NDF, ADF, cellulose and lignin). The pH variation and humidity in each substrate were determined, as well as the amino acidic composition. The fermentation effectiveness was tested by infrared spectroscopy. Changes in the physico-chemical and mineral characteristics occurred in the legumes studied with remarkable increase of CP and TP, and a decrease of NDF. In the enzymatic kinetics, there was interaction in every factor (P< 00.1) for the four legumes. Maximum endo âD1-4 glucanase and exo âD1-4 glucanase values were reached in Vigna unguiculata, with 18.10 and 12.71 IU/mL at 72h and 24 h, respectively. The amino acidic pattern totally differed in content for the final fermented products. The analysis with infrared spectroscopy in Vigna unguiculata showed differences in bands intensity (1660-1100cm-1), due to fermentation. The strain T. viride M5-2 allowed the development of a biologically feasible fermentation process with meals of legume grains under study to improve its nutritional value, allowing the inclusion of these formulations on diets of monogastric animals. Furthermore, the results show the possibility of obtaining an inoculum without the addition of other nutrients in the medium, which is very important for the process of scaling.
Key words: seasonal legumes, Trichoderma viride, fermentation, fungi.
RESUMEN
Para mejorar el valor nutritivo de leguminosas temporales mediante el procesamiento de sus granos con Trichoderma viride M5-2, se realizaron varios experimentos a escala de laboratorio con esta cepa mutante, sin adición de fuentes de nitrógeno ni minerales para distintas variantes de fermentación. Se utilizaron diseños completamente aleatorizados, con arreglo factorial 4 x 6 y tres repeticiones. Los factores variaron según la variante de análisis a utilizar y fueron los sustratos de Vigna unguiculata (vigna), Canavalia ensiformis (canavalia), Stizolobium niveum (mucuna) y Lablab purpureus (dólico), y los tiempos de fermentación (0, 24, 48,72, 96 y 120 h). Se realizaron muestreos cada 24 h para las determinaciones de actividad celulolítica (endo â1-4 glucanasa y exo â1-4 glucanasa), composición mineral (Ca, Mg, P, cenizas) y composición bromatológica (PB, PV, FND, FAD, celulosa y lignina). Se determinó la variación del pH y comportamiento de la humedad en cada sustrato, así como la composición aminoacídica y se comprobó la efectividad de la fermentación por FT-IR. Se produjeron cambios en las propiedades físico-químicas y minerales de las leguminosas estudiadas, con notable incremento de la PB, PV y disminución de la FND. En la cinética enzimática, para las cuatro leguminosas, hubo interacción en todos los factores (P < 00.1). Se alcanzaron valores máximos de enzimas endo âD1-4 glucanasa y exo âD1-4 glucanasa en Vigna unguiculata, de 18.10 y 12.71 UI/mL a las 72 h y 24 h, respectivamente. El patrón aminoacídico difirió totalmente en su contenido para los productos finales fermentados. El análisis por espectroscopía infrarroja en Vigna unguiculata mostró diferencias en la intensidad de las bandas (1660-1100 cm-1), debido a la fermentación. La cepa T. viride M5-2 permitió desarrollar un proceso fermentativo biológicamente factible con las harinas de granos de las leguminosas en estudio, al mejorar su valor nutritivo. Esto permitiría la incorporación de estas formulaciones en las dietas de animales monogástricos. Además, los resultados demuestran que es posible obtener un inoculo sin adición de otros nutrientes en el medio, aspecto de gran importancia para el proceso de escalado.
Palabras clave: leguminosas temporales, Trichoderma viride, fermentación, hongos.
INTRODUCTION
Seasonal legumes are species that adapt to a wide range of soil texture and fertility. They are nitrogen fixer plants that may be planted purely or associated with different soil rotation systems. From the nutritional point of view, their grains are characterized by a high protein and energy content, together with a considerable contribution of feeding fiber, vitamins and minerals.
The presence of toxic or anti-nutritional compounds on these legumes is the main nutritional limitation for their use in animal feeding, mainly in monogastric species, due to the morphological and physiological characteristics of their gastrointestinal tract. Findings of Diaz et al. (2003) and Martín-Cabrejas et al. (2008) about the content of antinutritional factors (ANFs) in these legumes showed several factors of different chemical nature and location in their grains like proteases inhibitors, lectins, phytates and tannins. However, a proper biotechnological process of their grains or meals, due to the action of Trichoderma viride M5-2, strain of a mutant fungi hyper-producer of cellulase and polyphenoloxidase enzymes, resistant to catabolic repression, could eliminate or reduce the presence of such antinutritional compounds and then increase the nutrients bioavailability in the gastrointestinal tract. The effectiveness of this fungus has been confirmed for degrading native and crystalline sugarcane bagasse cellulose with the cellulolitic complex it produces and segregates (Valiño et al. 2004). This would allow the increase of inclusion levels of Vigna unguiculata on diets for monogastric species and the obtaining of a superior productive response. Canavalia, dolicho and mucuna could be included as new sources of non-conventional feeds.
This biotechnological process (Zang y Lynd 2006, faria et al. 2008, Sipos et al. 2010 and Hurt et al. 2014) generates biochemical and structural modifications that allow to eliminate great part of these components with a consequent increase of the nutritive value of the resulting product. For that, the enzymatic activity is needed to carry out the hydrolysis of the â 1-4 glycosidic bonds, in a sequence operation and synergic action of the cellulase enzymes. They have different types of bonds due to the complex nature of the cellulose, which, besides, is included in a matrix of hemicellulose, pectin and lignin (Ericsson and Bermek, 2009). The mayor compound of hemicellulose is the mannan and contains sugars like mannose, galactose and glucose. The composition is variable and includes galactomannan and galactoglucomannan, and, in the case of pectin, the largest component is the galacturonic acid and includes ramnose, galactose, fucose and apiose (Willats et al. 2009). The composition of several lignocellulosic materials has been reported in the literature (Cianchetta et al. 2012, van Dyk and Pletschke 2012 and Hasunuma et al. 2013) and varies substantially depending on the source.
The objective of this study was to improve the nutritive value of the seasonal legumes Vigna unguiculata, Canavalia ensiformis, Stizolobium niveum and Lablab purpureus through processing their grains with Trichoderma viride M5-2, to obtain new products destined to animal feeding, as an alternative of conventional protein sources.
MATERIALS AND METHODS
Microorganism. A mutant strain of the lignocellulolitic fungus Trichoderma viride M5-2, belonging to the strain bank of the Institute of Animal Science, was used. This strain produces phenoloxidase and cellulase enzymes, has hydrolytic activity in highly fibrous substrates, assessed through fermentation in solid state (Valiño et al. 2004).
Fermentation substrates. Grains of seasonal legumes Vigna unguiculata var. Habana 82 (cowpea), Lablab purpureus (dolicho) Canavalia ensiformis (canavalia), and Stizolobium niveum (mucuna) were used.
Preparation of meals with legume grains. Legumes were sown in spring on a typical red ferralitic soil (Hernández et al. 1999), and harvested after completing the process of seed maturation, when 95% of pods were dried. After completing the process of threshing-benefit, the grains were sun-dried for two days until reaching humidity between 14 and 12%. Later, they were stored between 6 and 10 °C, with a relative humidity below 85%. For the fungal growth test and the fermentation experiment, legume beans were dried in an oven at 60 °C and ground in a hammer mill until reaching a particle size of 0.5 mm ± 0.2 mm.
Fermentation process. Erlenmeyer flasks of 500 mL were used, with 10 g of each type of legume grains. They were humidified with distilled water up to 70 % and no nutrients were added. The humid substrates were sterilized in an autoclave for 20 min. As inoculum, 1 cm2 of malt agar was used with the cultivated strain at 30 ºC for 7 days. The mixture was homogenized and the flasks were placed in an incubator at 30 ºC for 120 h. Samplings were performed every 24 hours, to conduct the corresponding chemical and enzymatic analysis.
Chemical and enzymatic analysis. An amount of 5g of the fermented solid material was taken at 0, 24, 48, 72, 96, and 120 h. A total of 45 mL of distilled water were added, and agitated in a sieve at 150 rpm for 30 min and, then, filtered to obtain the enzymatic extract, to which the pH was measured and the corresponding enzymatic analyses were conducted. The enzymes determinate were:
1. Endo 1, 4 ß glucanase (CMCase) that shows the hydrolytic activity over the carboxymetylcellulose.
2. Exo 1, 4 ß-D glucanase (PFase) or cellulase filter paper that shows hydrolytic activity in the presence of crystalline cellulose.
The activities of endo 1,4 ß- glucanase and exo 1,4 ß glucanase were determined, calculated and expressed in International Units per milliliters (IU/mL). This activity refers the glucose micromoles released per minute of reaction under the conditions of activity assay test (Mandels and Andreotti1976).
Bromatological analysis. Studied indicators were dry matter (DM), ash and crude protein (CP), according to AOAC (1995), true protein (TP) according to the method of Berstein (cited by Meir 1986), neutral detergent fiber (NDF), acid detergent fiber (ADF), lignin (LIG), and cellulose (CEL) according to Goering and Van Soest et al. (1991). Ca, P and Mg were measured by atomic absorption.
Amino acid analysis. For the analysis of the amino acidic pattern, all samples were previously degreased by the extraction under reflux in petroleum ether. Subsequently, the samples were treated with HCl 6N under reflux conditions, at 110 °C, in order to cleave the peptide chain and guarantee the presence of amino acids as free molecules in solution to be analyzed. The hydrolyzed materials were processed in the amino acid analyzer ALPHA PLUS II. The general requirements for developing standard operating procedures (PNO AC. 01. 001. 94 in Spanish) and for determining total amino acids (PNO ID. 05. 002. 95 in Spanish) were considered. Three samples were analyzed for Vigna unguiculata.
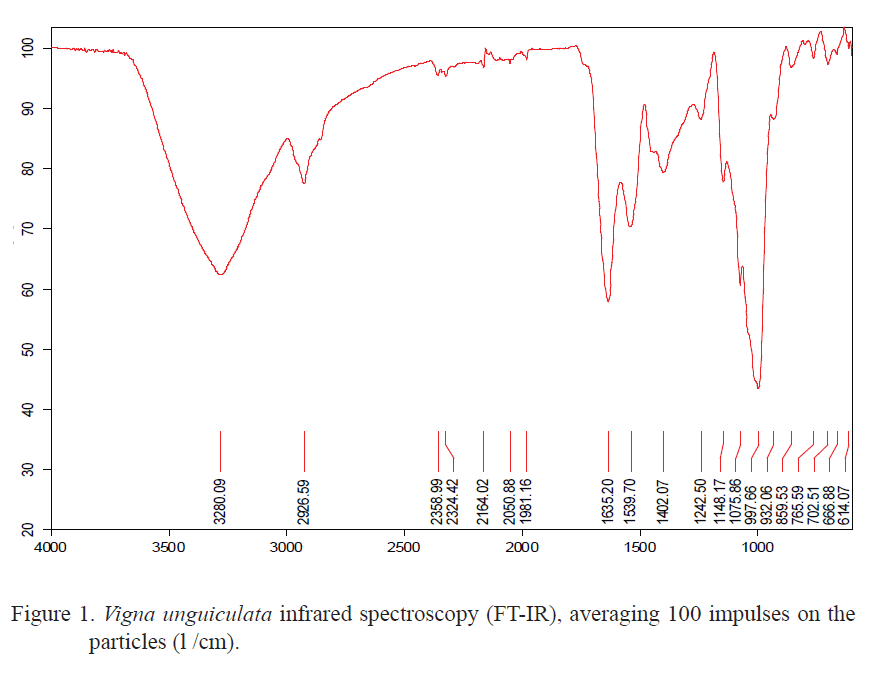
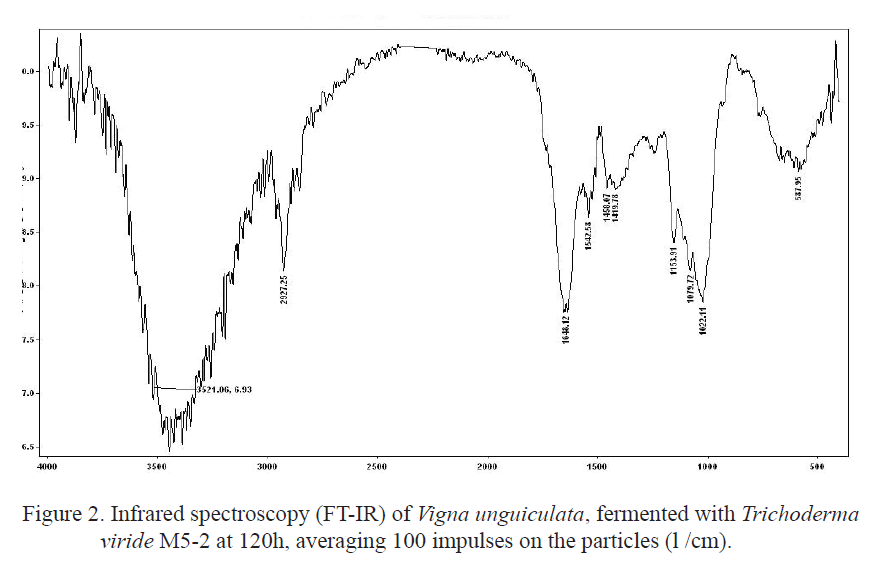
Analysis using Fourier Transformed-Infrared Spectroscopy (FT-IR). The solid fermented samples were selected for FT-IR analysis, obtaining the maximum cellulolitic activity. The equipment used was an ATI Mattson Genesis Series FT-IR spectrometer, with a special device for solid samples, adjusted to total attenuated reflectance in pieces of potassium bromide (KBr). This allowed measuring the IR spectrum in the solid samples without previous preparation. The spectra were obtained in form of transmittance, in a wave longitude range of 4,000-600 cm-1, averaging 100 impulses over the particles of legume grain meal (ë/cm).
Statistical analysis. A completely randomized design was used, with a 4x6 factorial arrangement and three repetitions of four substrates [Vigna unguiculata (cowpea), Canavalia ensiformis (canavalia), Stizolobium niveum (mucuna), and Lablab purpureus (dolicho)] and six sampling times (0, 24, 72, 48, 96, 120 h) for enzymes, pH and DM. For minerals and bromatological composition, two sampling times were used (0 and 120 h). Each Erlenmeyer was considered as an experimental unit. For enzymatic and bromatological analysis, the statistical package INFOSTAT, version 1.0 (Di Rienzo et al. 2012), from the National University of Cordova, Argentina was used. The differences between means were established for the necessary cases, according to Duncan (1955).
RESULTS AND DISCUSSION
Table 1shows, the Trichoderma viride M5 -2 fungus grew in the four legumes studied without addition of nutrients and only with the appropriate humidity level for their development. The observed differences in growth were confirmed after the sporulation of the fungus. Mycelial growth started after 24 hours of inoculation in cowpea and dolicho substrates. After 72 hours of fermentation, both substrates were completely covered in spores, which may promote greater cellulolitic activity. Besides, this factor makes the fungus more competitive for reducing the antinutritional factors during fermentation. The particle size also favored greater attack of the fungus, when to contact surface increased (Chundawat et al. 2007, Alvira et al. 2010 and Fitzpatrick et al. 2010). According to Savón et al. (2002), the reduction to particle size or in volume decreases the flow speed of digesta in the gastrointestinal tract of monogastric animals and allows a higher action of digestive enzymes and, consequently, a better utilization of nutrients by the animal.
The growth in Canavalia ensiformes started after 48 h, and was completely covered with bright green sporulation at 72 h. However, mucuna growth was different, with a pink coloration in the spores. Further studies should analyze the possible relation of morphological variations, which is a result from compounds contained in the mucuna grain that favor these changes in color.
Out of this result, an experiment in solid state fermentation was performed to determine the changes in chemical, physical and enzymatic indicators and assess the potential of meals of tropical legume grains for feeding monogastric species.
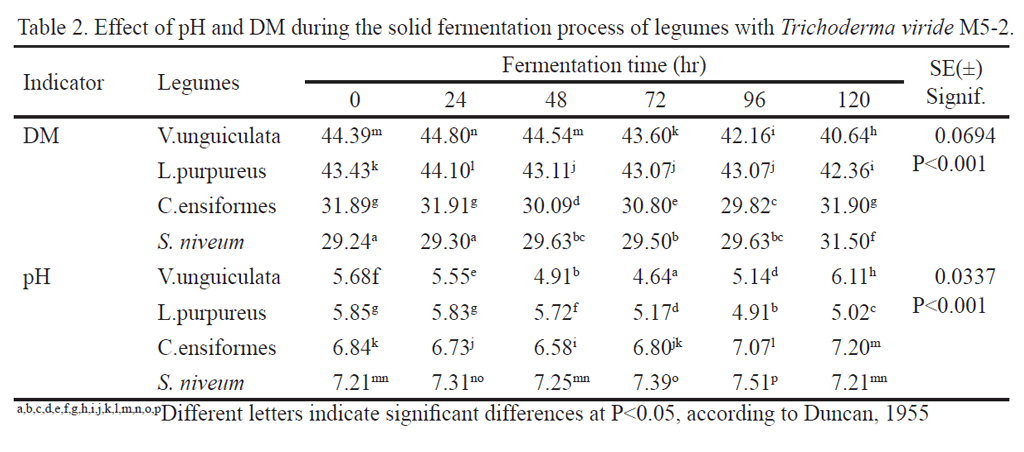
Table 2 shows the fermentation process of the four legumes used in pH and DM, where interaction of factors in study occurred. Values of pH under 6 (P < 0.001) were observed for Lablab purpureus and Vigna unguiculata. This coincides with the action range of cellulolitic enzymes in highly fibrous substrates like sugarcane because it is known that the initial optimum pH for hydrolytic action of cellulases are between 5 and 6 (Mandels et al. 1982 and Leuchmann, 1996) depending on the substrate to be fermented and growth temperature of each microorganism, although the strain used is resistant to catabolic repression and cellulolitic action range is between 5 and 7 (Valiño et al. 2004 and Valiño et al. 2011). In the case of Canavalia ensiformes and Stizolobium niveum, pH was above 6, up to values of 7.39. This could be related to groups á amino that, when dissolved in water, are ionized and a proton of amino group that could act as base is removed (Bover-Cid et al. 2014), although this condition did not affect the growth of the fungus.
There are differences in the performance of humidity in fermentation times studied (table 2). The dry matter decreased in 3.45 percentage units in the grain meal of Vigna unguiculata. However, Lablab purpureus decreased only 1.12%, a feature to be considered for escalation processes. Around 30% remained in other legumes. The humidity level for fermentation should not exceed the water retention level, taken as the upper limit to 80% of humidity. Moreover, if the humidity content in the substrate is very high, oxygen level and gas volume between the particles decrease. This decreases the interchanging effect and increases the risk of bacterial contamination (Pandey 2001 and Dustet and Izquierdo 2004). Likewise, humidity determines physical and chemical changes like composition of soluble carbohydrates, amount of phytate and alkaloids, changes modifying the nutritional value and, consequently, the character of legumes as functional food. Although humidity did not adversely affect the development of fermentation in this study, it is recommended to determine it for each species and, probably, for each strain, depending on the productive process.
In fermentation processes, the hydrolysis of antinutritional compounds depends on the type of legume and the type of fermentation process. Biotransformations occurring to innocuous compounds are the result of the combined effect of the action of internal enzymes within grains and of microorganisms responsible for fermentation (Seena et al. 2006 and Sreerama et al. 2012).
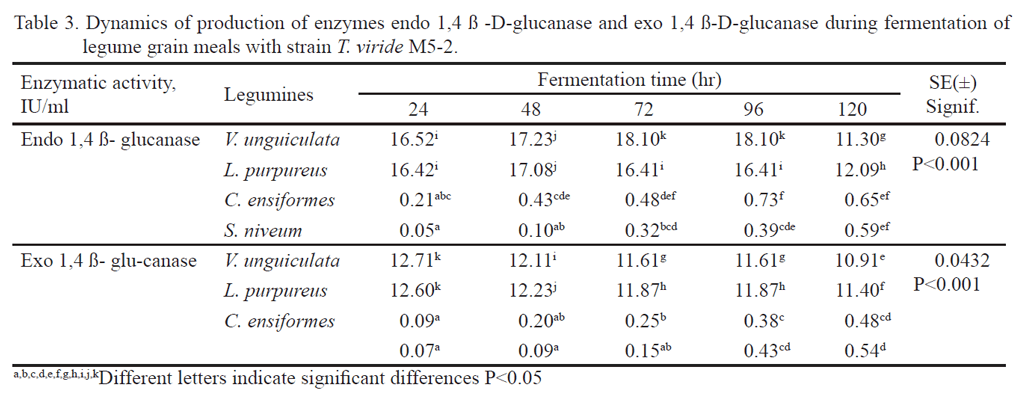
The analysis of the enzyme production dynamics showed that there was interaction between the factors studied (P <0.001) for the enzyme activities (endo â1-4 glucanase and exo â1-4 glucanase). Also, there was higher cellulolitic activity in Vigna unguiculata in a shorter fermentation time with Trichoderma viride strain M5-2. The cellulolitic activity began to increase between 0-24 hours in correspondence to mycelial growth. In Vigna unguiculata, the highest values were obtained at 72 and 96 h with 18.10 IU/mL of CMCase and 12.71 IU/mL PFasa at 24h. In Lablab purpureus, maximum production of CMCase was obtained at 48 h, with 17.08 IU/mL, and at 24 h for PFasa with 12.6 IU / mL with T. viride M5-2, and were in the range between 10 and 18 IU/mL (table 3). However, for Canavalia ensiformes and Stizolobium niveum, these enzymes values were very low and do not correspond to the results of the fungus growth during fermentation.
This performance is due to, according to Andersen et al. 2008, the relation between the two enzymes and the individual sum of the hydrolytic action on the substrate, which favors the synergy degree that may increase the improvement percentage of the enzymatic activity (Gottschalk et al. 2010 and Yoon 2014). The synergy degree is the quantification of the capacity of two or more enzymes for their action on the substrate. In this experiment, neither the enzyme â glucosidase nor xylanase were determined, only those with higher weight on hydrolytic activity of the crystalline and amorphous cellulase were quantified. In agreement with Andersen et al. (2008), this synergy degree could be based on the product formation, substrate conversion or reaction degree. This way, information about the substrate degradation and action mechanisms can be given (Gao et al. 2011 and Moscon et al. 2014).
To include grain meals, fermented in the diets of monogastric animals, it is necessary to characterize the fiber fraction. This includes the chemical composition and the structure of their cell walls, origin and nature, which helps to determine the quality of fibrous foods and predicts their effects on gastrointestinal and metabolic functions of the animal organism. Table 4 shows the effect of the fermentation process on the fibrous and protein content of grain meals of the studied temporary legumes. The short adaptation period and good growth of this strain in the four legumes demonstrated their valuable potential in the biotransformation of substrates, because protein indicators increased and NDF decreased. In the fermented Vigna unguiculata, there was an increase of CP in 6.27 percentage units and of TP in 5.11 percentage units, with the highest decrease of NDF (12.73%) during the process regarding the rest of the legume. However, the value increased for the other fiber (ADF and lignin). Cellulose showed no changes, taking into account the main effects of legumes and fermentation time with cellulolitic action.
These results coincide with studies of Diaz et al. (2003), Savon et al. (2000) and Savón (2005) where these unconventional unfermented legumes showed higher content of insoluble fiber (hemicellulose and lignin), from 303 to 399 mg kg-1 DM, regarding soluble fiber (pectin, gums, mucilage and polysaccharides). Cellulose showed high variability, Vigna unguiculata had the lowest content of the insoluble fraction, while Stizolobium niveum showed the highest contents. Therefore, the changes that occurred involve changes in the physical and chemical properties of these legumes, so fermentation can be considered as an effective method to improve its nutritional value, depending on the species and on the conditions under which the procedure is conducted, as well as age and other related environmental conditions (season and temperature) that can modify the nutritional value of fibrous fractions in grain meals.
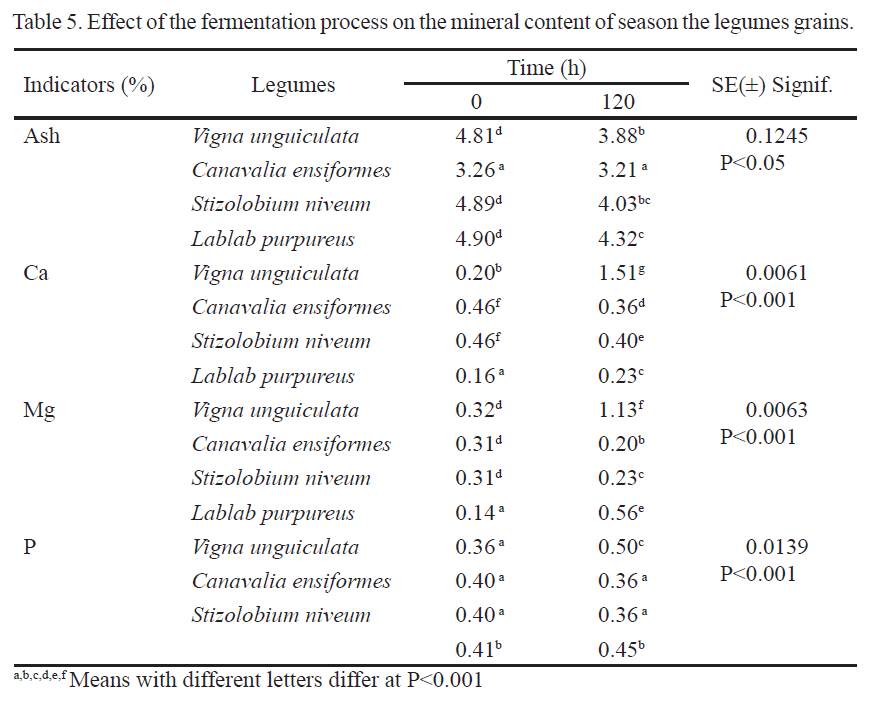
During the process of solid state fermentation, there were increases of Ca, Mg and P in Vigna unguiculata and Lablab purpureus, even when no mineral or nitrogen source was added (table 5). On the other hand, ash diminished, probably due to the volatilization process of compounds like ammonia during fermentation, very significant aspect according to the composition of these plant materials. On the contrary, these minerals diminished in Canavalia ensiformes and Stizolobium niveum, apparently associated to the own nature of these substrates, which have higher content of antinutritional factors and to the growth process of the used fungi and its transformation into TP. Regarding mineral availability, it has been demonstrated that, after a germination process, availability of phosphorous, potassium, magnesium, zinc, and copper increases as a direct consequence of the activation of phytase enzyme, which causes a decrease of inositols phosphates hexa and penta phosphorilates to less phosphorilated shapes (Ragab et al. 2004, Adebowale et al. 2005 and Belane and Dakora 2011).
From the results achieved in the cellulolitic activity, CP, TP and NDF of Vigna unguiculata, amino acid composition and molecular structure of this variety were studied. Besides, the variety Habana 82 has the highest agro-productive potential, according to Díaz et al. (2002).
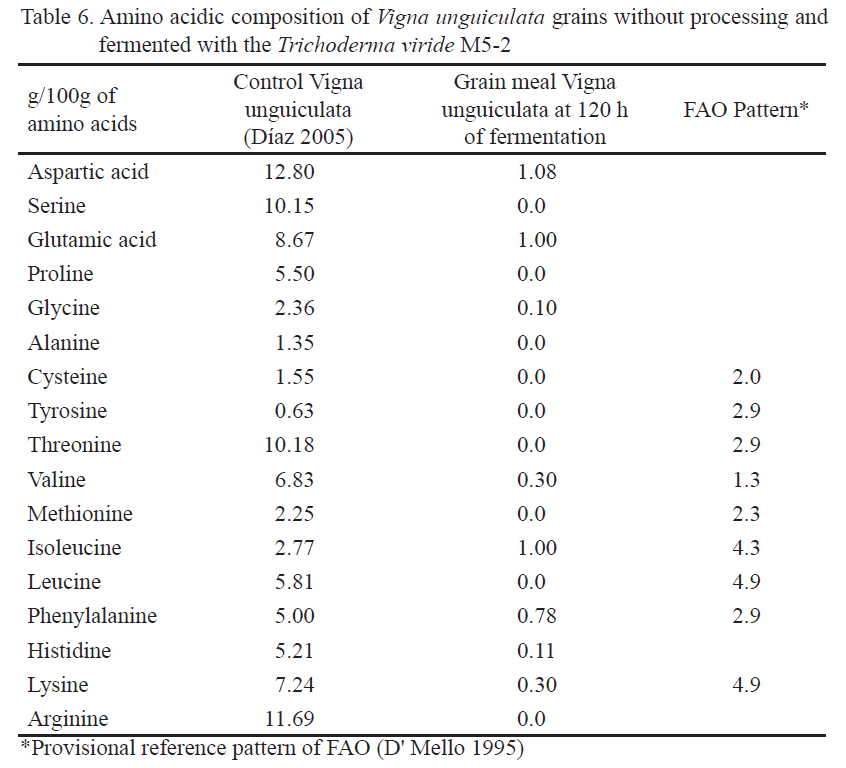
Changes of amino acidic patterns of Vigna unguiculata were observed during the fermentation. The essential amino acids (threonine, leucine, and arginine) and the non essential ones (serine, glycine, alanine, cysteine, and thyroxin) had the highest variation, because they were not present at the end of the fermentation (table 6). The rest diminished considerably, which could be related to the activities of the cellulolitic enzymes, endo 1, 4 ß- glucanase and exo 1, 4 ß glucanase, excreted by the fungi for degrading the substrate. These enzymes can join to Vigna unguiculata through its active site or aminoacids sequences (carbohydrates bond module, CBM) involved in recognizing bonds of polysaccharides, allowing its union with the substrate while hydrolysis of these chains occurs (Boraston et al. 2004), in function of the increase in the unicellular protein, contrary to that occurring with germination. However, this legume, without processing, showed a good amino acidic balance, with concentrations of essential amino acids, similar or superior to those established as reference pattern by FAO (Díaz et al. 2002 and Díaz et al. 2007). Non essential amino acids explain the highest variability percentage among species and other legume varieties related to this study, and not in the fermentative process.
It is necessary to deepen on the effect of legume fermentation by fungi producers of cellulase on the amino acidic composition, to obtain more information about nutritional value, and validate its nature as functional food acquired by the grains of these legumes, after going through this biological process. Further studies on protein fractioning and proteolytic activity during fermentation should be performed It is important to analyze the molecular structure of the variety Vigna unguiculata. For this purpose, the infrared spectroscopy was used, which is one of the most commonly used techniques to detect the presence of a compound of functional groups. Although the infrared spectrum characterizes each compound, certain atomic clusters always result in bands in certain range of frequencies, apart from the nature of the rest of the molecule.
Figure 1 and 2 shows the corresponding spectrum of the fermented and unprocessed Vigna unguiculata samples. The solid samples, in general, have well defined bands and of good resolution. The difference between the samples of unprocessed Vigna unguiculata and fermented Vigna unguiculata is evident, in the range between 2.000 and 1.400 cm-1. In this last, the bands are more opened. In the range 3.270-3.420 cm-1, there is an intense band of valence vibration of the O-H y OH. This band results from the polymeric association of the OH group of carbohydrates. In 2,930 cm-1, the band has mid intensity, according to the valence vibration of the C-H y CH bond of alkyl groups from the same carbohydrates chain.
The characteristic band of carbonyl groups present in amides of 1665-1640 cm-1, intense band of valence vibration from the C=O y C=O carbonyl group, appears slightly unfolded in two, so the presence of amino acids or proteins in the samples is inferred. This band has such intensity that it is unlikely to associate it with any other bond, which could be explained by legume proteins. In this case, Vigna unguiculata has an inferior protein quality to that of casein, but very superior to that of dolicho (Savón 2005 and Díaz et al. 2007). The fermentation process of Vigna unguiculata meal demonstrated the effectiveness of the analysis developed in the biological indicators. However, between 1.445 and 1.390 cm-1, the formation bands of the OH in carbohydrates have low intensity, unlike the range between 1.161 and 1.025 cm-1 that are very intense and correspond to valence vibration of the C-O bond, present in carbohydrates, esters and phenols. These data obtained by FT-IR in fermented Vigna unguiculata do not coincide with those obtained in the bromatological analysis of these samples by T. viride M5-2 when observing a lignin concentration of 2.73% and 2.4% of the cellulose on DM basis. This suggests that the lingocellulosic composition of plant materials may vary, according to the methods of analysis (Valiño et al. 2004 and Foyle et al. 2007), considering also the differences confirmed in these indicators during the harvest periods of the crops (Himmel et al.2007).
The technique demonstrated that, during the solid state fermentation with the strain of conidial fungus in Vigna unguiculata, oxidations of aromatic structures of lignin occur, which lead to the increase of O-H phenolic groups. Their rupture leads to the formation of alkene, carbonyl and carboxyl groups, as well as alcohol, carbonyl and carboxyl bonds, derived from biodegradation of carbohydrates that cannot be detected by other means.
The strain T. viride M5-2 allowed the development of a biologically feasible fermentation process with grain meals of legumes under study, when improving their nutritional value. This allows the inclusion of these diet formulations for monogastric animals. Furthermore, the results demonstrated the possibility of obtaining an inoculum without addition of other nutrients to the medium, which is important to the scaling of these fermentation processes.
BIBLIOGRAPHY
Adebowale Y. A., Adeyemi A. & Oshodi A. A. 2005. ‘‘Variability in the physicochemical, nutritional and antinutritional attributes of six Mucuna species’’. Food Chemistry, 89 (1), pp. 37–48.
Adebowale Y. A., Adeyemi I. A., Oshodi A. A. & Niranjan K. 2007. ‘‘Isolation, fractionation and characterisation of proteins from Mucuna bean’’. Food Chemistry, 104 (1), pp. 287–299.
Alvira P., Tomás-Pejó E., Ballesteros M. & Negro M. J. 2010. ‘‘Pretreatment technologies for an efficient bioethanol production process based on enzymatic hydrolysis: a review’’. Bioresource technology, 101 (13), pp. 4851–4861.
Andersen N., Johansen K. S., Michelsen M., Stenby E. H., Krogh K. B. & Olsson L. 2008. ‘‘Hydrolysis of cellulose using mono-component enzymes shows synergy during hydrolysis of phosphoric acid swollen cellulose (PASC), but competition on Avicel’’. Enzyme and Microbial Technology, 42 (4), pp. 362–370.
AOAC. 1995. Official Methods of Analysis. 16th ed., Washington: Assoc. Off. Agric. Chem.
Belane A. K. & Dakora F. D. 2011. ‘‘Levels of nutritionally-important trace elements and macronutrients in edible leaves and grain of 27 nodulated cowpea (Vigna unguiculata L. Walp.) genotypes grown in the Upper West Region of Ghana’’. Food Chemistry, 125 (1), pp. 99–105.
Boraston A. B., Bolam D. N., Gilbert H. J. & Davies G. J. 2004. ‘‘Carbohydrate-binding modules: fine-tuning polysaccharide recognition’’. Biochemical Journal, 382 (3), pp. 769–781.
Bover-Cid S., Latorre-Moratalla M. L., Veciana-Nogués M. T. & Vidal-Carou M. C. 2014. ‘‘Processing contaminants: biogenic amines’’. In: Encyclopedia of Food Safety, vol. 2, pp. 381–391.
Chundawat S. P., Venkatesh B. & Dale B. E. 2007. ‘‘Effect of particle size based separation of milled corn stover on AFEX pretreatment and enzymatic digestibility’’. Biotechnology and bioengineering, 96 (2), pp. 219–231.
Cianchetta S., Galletti S., Burzi P. L. & Cerato C. 2012. ‘‘Hydrolytic potential of Trichoderma sp. strains evaluated by microplate-based screening followed by switchgrass saccharification’’. Enzyme and microbial technology, 50 (6), pp. 304–310.
Díaz M. F., González A., Padilla C. & Curbelo F. 2002. ‘‘Caracterización bromatológica de granos y forrajes de las leguminosas temporales Canavalia ensiformis, Lablab purpureus y Stizolobium niveum sembradas a finales de la estación lluviosa’’. Rev. cubana Cienc. agríc, 36 (4), p. 409.
Diaz M. F., Martin-Cabrejas M. A., Gonzalez A., Torres V. & Noda A. 2007. ‘‘Biotransformation of Vigna unguiculata during the germination process’’. Cuban Journal of Agricultural Science, 41 (2), pp. 161–165.
Díaz M. F., Padilla C., Torres V., González A. & Noda A. 2003. ‘‘Caracterización bromatológica de especies y variedades de leguminosas temporales con posibilidades en la alimentación animal’’. Cuban J. Agric. Sci, 37 (4), Available: <http://repositoriodigital.academica.mx/jspui/handle/987654321/386998>, [Accessed: November 3, 2014].
Di Rienzo J. A., Casanoves F., Balzarini M. G., González L. & Robledo C. W. 2012. InfoStat. version 2012, [Windows], Universidad Nacional de Córdoba, Argentina: Grupo InfoStat, Available: <http://www.infostat.com.ar/> .
D’Mello J. P. F. & Devendra C. 1995. ‘‘Leguminous leaf meals in non-ruminant nutrition’’. In: Tropical legumes in animal nutrition, Wallinford, UK: CAB International, pp. 247–282, Available: <http://www.cabdirect.org/abstracts/19951408994.html>, [Accessed: November 3, 2014].
Dustet M. J. C. & Izquierdo K. E. 2004. ‘‘Aplicación de balances de masa y energía al proceso de fermentación en estado sólido de bagazo de caña de azúcar con Aspergillus niger’’. Biotecnología Aplicada, 21 (2), pp. 85–91.
Eriksson K.-E. & Bermek H. 2009. ‘‘Lignin, lignocelluloses, ligninase’’. Appl. Microbiol. Industrial, pp. 373–384.
Faria L. F., Kolling D., Camassola M., Dillon A. J. P. & Ramos L. P. 2008. ‘‘Comparison of Penicillium echinulatum and Trichoderma reesei cellulases in relation to their activity against various cellulosic substrates’’. Bioresource Technology, 99 (5), pp. 1417–1424.
FitzPatrick M., Champagne P., Cunningham M. F. & Whitney R. A. 2010. ‘‘A biorefinery processing perspective: treatment of lignocellulosic materials for the production of value-added products’’. Bioresource technology, 101 (23), pp. 8915–8922.
Foyle T., Jennings L. & Mulcahy P. 2007. ‘‘Compositional analysis of lignocellulosic materials: Evaluation of methods used for sugar analysis of waste paper and straw’’. Bioresource Technology, 98 (16), pp. 3026–3036.
Gao D., Uppugundla N., Chundawat S. P., Yu X., Hermanson S., Gowda K., Brumm P., Mead D., Balan V. & Dale B. E. 2011. ‘‘Hemicellulases and auxiliary enzymes for improved conversion of lignocellulosic biomass to monosaccharides’’. Biotechnology for biofuels, 4 (1), p. 1.
Goering H. & Van Soest J. P. 1970. Forage fiber analysis. (ser. Agricultural Hand book, no. ser. 379), Washington, USA: US Dept. Agriculture.
Gottschalk L. M., Oliviera R. A. & Da Silva Bon E. . 2010. ‘‘Cellulase, Xylanases, βGlucanasa and acid ferulic, acid estearease produced by Trichoderma and Aspergillus act synergistically in the hydrolysis of sugar cane bagasse’’. Bioresource Technol, 51, p. 72.
Hasunuma T., Okazaki F., Okai N., Hara K. Y., Ishii J. & Kondo A. 2013. ‘‘A review of enzymes and microbes for lignocellulosic biorefinery and the possibility of their application to consolidated bioprocessing technology’’. Bioresource technology, 135, pp. 513–522.
Himmel M. E., Ding S.-Y., Johnson D. K., Adney W. S., Nimlos M. R., Brady J. W. & Foust T. D. 2007. ‘‘Biomass recalcitrance: engineering plants and enzymes for biofuels production’’. science, 315 (5813), pp. 804–807.
Hur S. J., Lee S. Y., Kim Y.-C., Choi I. & Kim G.-B. 2014. ‘‘Effect of fermentation on the antioxidant activity in plant-based foods’’. Food chemistry, 160, pp. 346–356.
Leuchtmann A., Petrini O. & Samuels G. J. 1996. ‘‘Isozyme subgroups in Trichoderma section Longibrachiatum’’. Mycologia, 88, pp. 384–394.
Mandels, M. 1982. ‘‘Cellulases’’. Anual Reports on Fermentation Process, 5, p. 78.
Mandels M., Andreotti R. E. & Roche C. 1976. ‘‘Measurement of scarifying cellulose’’. In: Biotechnol. Bioeng. Symp, vol. 6, p. 1471.
Martín-Cabrejas M. A., Díaz M. F., Aguilera Y., Benítez V., Mollá E. & Esteban R. M. 2008. ‘‘Influence of germination on the soluble carbohydrates and dietary fibre fractions in non-conventional legumes’’. Food Chemistry, 107 (3), pp. 1045–1052.
Meir H. 1986. ‘‘Laborpraktikure. Tierernahurungund, futtermittelm für Tierproduzenten’’. Biotech. Tech, 2 (1).
Moscon J. M., Priamo W. L., Bilibio D., Silva J. R., Souza M., Lunelli F., Foletto E. L., Kuhn R. C., Cancelier A. & Mazutti M. A. 2014. ‘‘Comparison of conventional and alternative technologies for the enzymatic hydrolysis of rice hulls to obtain fermentable sugars’’. Biocatalysis and Agricultural Biotechnology, 3 (4), pp. 149–154.
Pandey A. 1991. ‘‘Aspects of fermenter design for solid-state fermentations’’. Process Biochemistry, 26 (6), pp. 355–361.
Ragab D. M., Babiker E. E. & Eltinay A. H. 2004. ‘‘Fractionation, solubility and functional properties of cowpea (Vigna unguiculata) proteins as affected by pH and/or salt concentration’’. Food chemistry, 84 (2), pp. 207–212.
Rangel A., Saraiva K., Schwengber P., Narciso M. S., Domont G. B., Ferreira S. T. & Pedrosa C. 2004. ‘‘Biological evaluation of a protein isolate from cowpea (Vigna unguiculata) seeds’’. Food Chemistry, 87 (4), pp. 491–499.
Savón L. 2002. ‘‘Alimentos altos en fibra para especies monogástricas. Caracterización de la matriz fibrosa y sus efectos en la fisiología digestiva’’. Revista Cubana de Ciencia Agrícola, 36 (2), pp. 91–102.
Savón L. 2005. ‘‘Alimentos fibrosos tropicales y su efecto en la fisiología digestiva de especies monogástricas’’. Rev. Cubana Cienc. Agríc, 39, p. 475.
Savón L., Leyva O. & González T. 2000. ‘‘Caracterización físico química de la fracción fibrosa de cinco variedades de harina de forraje de Vigna unguiculata para especies monogástricas’’. Rev. Cubana Cienc. Agríc, 34, p. 137.
Seena S., Sridhar K. R. & Jung K. 2006. ‘‘Nutritional and antinutritional evaluation of raw and processed seeds of a wild legume, Canavalia cathartica of coastal sand dunes of India’’. Food chemistry, 92 (3), pp. 465–472.
Sipos B., Benk\Ho Z., Dienes D., Réczey K., Viikari L. & Siika-aho M. 2010. ‘‘Characterisation of specific activities and hydrolytic properties of cell-wall-degrading enzymes produced by Trichoderma reesei Rut C30 on different carbon sources’’. Applied biochemistry and biotechnology, 161 (1-8), pp. 347–364.
Sreerama Y. N., Sashikala V. B., Pratape V. M. & Singh V. 2012. ‘‘Nutrients and antinutrients in cowpea and horse gram flours in comparison to chickpea flour: Evaluation of their flour functionality’’. Food Chemistry, 131 (2), pp. 462–468.
Valino E. C., Elías A., Torres V., Carrasco T., Albelo N. & others. 2004. ‘‘Improvement of sugarcane bagasse composition by the strain Trichoderma viride M5-2 in a solid-state fermentation bioreactor.’’. Cuban Journal of Agricultural Science, 38 (2), pp. 143–150.
Van Dyk J. S. & Pletschke B. I. 2012. ‘‘A review of lignocellulose bioconversion using enzymatic hydrolysis and synergistic cooperation between enzymes—factors affecting enzymes, conversion and synergy’’. Biotechnology advances, 30 (6), pp. 1458–1480.
Willats W. G., Knox J. P. & Mikkelsen J. D. 2009. ‘‘Pectin: new insights into an old polymer are starting to gel’’. Trends in Food Science & Technology, 17 (3), pp. 97–104.
Wilson D. B. 2011. ‘‘Microbial diversity of cellulose hydrolysis’’. Current opinion in microbiology, 14 (3), pp. 259–263.
Yoon L. W., Ang T. N., Ngoh G. C. & Chua A. S. M. 2014. ‘‘Fungal solid-state fermentation and various methods of enhancement in cellulase production’’. Biomass and Bioenergy, 67, pp. 319–338.
Zhang Y.-H. P. & Lynd L. R. 2006. ‘‘A functionally based model for hydrolysis of cellulose by fungal cellulase’’. Biotechnology and Bioengineering, 94 (5), pp. 888–898.
Received: November 3, 2014
Accepted: December 1, 2014
Elaine Valiño, Instituto de Ciencia Animal, Apartado Postal 24, San José de las Lajas, Mayabeque, Cuba. Email: evalino@ica.co.cu


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}