










 Custom services
Custom services text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroduction
The use of tropical foliages in rabbit feeding is a subject of interest for countries in which conventional systems of rabbit production (McNitt et al. 2000) may not be sustainable if locally available foliar sources are not used (Dihigo et al. 2001, 2008, Phinmasan et al. 2004, Pok Samkol et al. 2005, Nieves 2008 and Nieves et al. 2009 a, b). Moringa (Moringa oleifera) is one of the tropical foliages that could be of interest to feed rabbits (Pérez et al. 2010). Moringa is a plant species whose foliage is characterized by having an appreciable protein level and, in turn, low cell wall content. This foliage usually has high in vitro ileal digestibility of N (Ly et al.2001 and Mireles et al. 2017).
The yields of moringa foliage, as well as its chemical composition, have motivated the study of its nutritional value in rabbits in different tropics regions, particularly when the food is supplied in granular form or in multi-blocks (Odeyinka et al. 2008, Nuhu 2010, Rodríguez 2010, Adenji and Lawal 2012 and Vivas 2014) and not as meal (Caro et al. 2018).
The gastrointestinal tract of rabbits has particularities that notably differentiate it from other farm animals. Its characteristics allow through the caecotrophy, the successful extraction of valuable nutrients from the forages that are supplied (Lebas et al. 1986 y Carabaño and Piquer 1998). From the morphological point of view, it has been found that the different digestive segments grow at different speeds, between three and seven weeks of age (Lebas and Laplace 1972). From the point of view of the influence of food ingested by the rabbit, different characteristics of the macroarchitecture of the gastrointestinal tract have been associated with feeding strategies and with the yield of the carcasses (Gidenne et al. 2012), so the traits of the digestive organs may also have a practical interest.
Under feeding conditions with tropical forages, there is little information about the interdependence between this type of rabbit feeding and the morphological characteristics of the alimentary canal of animals, especially when rabbits are fed with moringa foliage (Odeyinka et al. 2008, Dougnon et al. 2012 and Odetola et al. 2012).The objective of this study was to determine if there are changes in the gastrointestinal macroarchitecture of rabbits fed with varying levels of moringa forage meal in the diet.
Materials and Methods
The research was developed jointly in the Instituto de Ciencia Animal and in a non-state farm, of provincial reference, located in Madruga municipality, Mayabeque province, between June and August 2012.
The tree species Moringa oleifera accession Supergenius (India) was evaluated. This forage was obtained in the forage area from the Instituto de Ciencia Animal. It was established on typical red ferralitic soil, with rapid desiccation and uniform profile (Hernández et al. 1999).
Experimental procedure. In the Estación Experimental de Pastos y Forrajes "Miguel Sistach Naya", belonging to the Instituto de Ciencia Animal, the cultivation of M. oleifera var. Supergenius was established. The sowing was manually done, at a density of 100 000 plants/ha in a circumscribed area of 200 m2. For the forage processing, the whole plant was sampled and a second cut was carried out about 60 d after the previous cut. The cut of this forage was manually made, at a height of 30 cm above soil level. The process of drying the material was done in the shade for a period of five days, in order to reduce the humidity.
To facilitate the drying of this material, as well as undesired fermentative processes, it was preceded to turning three times a day with a rake, taking into account a bedding height less than 30 cm. Then, it was reduced to a 3 mm particle size, in a hammer mill and the meal was stored in 50 kg jute sacks until use.
Samples were taken from six sacks, previously homogenized. In order to obtain a representative mixture of approximately 1 kg, the contents of the sack were spread on a clean and flat surface for random sampling. The particle size was reduced to 1 mm in a hammer mill and then passed through a sieve of equal size to ensure a uniform size. The mixture was packed in glass bottles with a hermetic seal and stored at room temperature until further analysis (Herrera et al. 1986). The composition of the material under study is shown in table 1.
Animals and experimental diets. Twenty-four White New Zealand male rabbits of 45 d of age and initial live weight of ± 885 g were used. The animals were allocated in individual galvanized wire metal cages, equipped with a linear feeder and an automatic nipple drinker. The total duration of the experiment was 45 d.
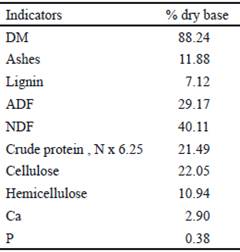
The composition of the experimental diets is shown in table 2. The diets were formulated according to the nutritional needs of protein and energy, established by Machado et al. (2011). They were isoproteic and isoenergetic. The treatments consisted on a control diet and the inclusion of 15 % and 30 % moringa forage meal. The diets were supplied as meal. During the experimental period the animals had food and water ad libitum. The food was offered in two portions (8:00 a.m. and 6:00 p.m.) daily.
Table 2 Chemical composition of experimental diets (percent in dry base)
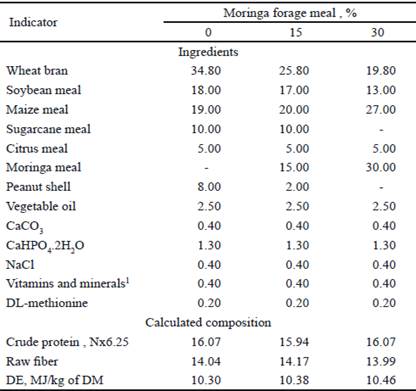
1 Each kg contains: vitamin A, 12 000 UI; vitamin D3, 2000 UI; vitamin B2,4160 UI, niacin, 16 700 UI, pantothenic acid , 8200 UI; vitamin B6, 3420 UI; folic acid, 0.98 g; vitamin B12, 16 mg; vitamin K, 1560 UI; vitamin E, 16 g; BHT, 8.50 g; cobalt, 0.75 g, copper; 3.50 g, iron, 9.86 g; manganese, 6.52 g; sodium, 0.87 g; zinc, 42.4 g; selenium, 6.60
Experimental procedure. The animals were slaughtered at the end of the period with 90 d of age and average weight ± 2.0 kg. After the laparatomy, the digestive tract was ligated in different places to avoid movements of digesta and four segments were isolated: stomach, small intestine, caecum and colon/ rectum. The organs were weighed full and empty. The difference between both weights was considered as the content of fresh digesta.
Morphometric indicators. The fresh weight of the gastrointestinal tract and each digestive organ was expressed as weight relative to live weight (g/kg) and percentage contribution of the digestive organs with respect to the total weight of the gastrointestinal tract (GIT), in %. The length of the intestinal segments was expressed relative to live weight (cm/kg), while the linear density was determined as relative weight of the intestines between their relative lengths (g/cm).
The digestive content of the stomach and caecal was expressed as relative weight to liveweight (g/kg) as well as the percentage contribution of the stomach and caecum with respect to the total weight of the GIT (%).
Experimental design and statistical analysis. A completely randomized design with three treatments and eight repetitions was used. The animal represented the experimental unit. The data of the evaluated indices were processed using the analysis of variance technique according to a simple classification, or regression (Steel et al. 1997). In the necessary cases, the means were compared by the Tukey test. Statistical packages of Minitab (2018) and InfoStat version 2012 (Di Rienzo et al. 2012) were used.
Results and Discussion
The data related to the weight of the full and empty digestive organs are shown in tables 3 and 4. In general, the preponderant role-played by the stomach and the caecum corresponded to their percentage contribution to the weight of the entire tract, which is already known (Lebas and Laplace 1972 and Carabaño and Piquer 1998).
The percentage contribution of the full stomach differed between treatments (table 3). The obtained results are comparable with those published by Diz (2013), who evaluated the inclusion of 20, 30 and 40 % of moringa forage meal in a commercial concentrate. In contrast, Dihigo et al. (2001) reported a decrease in the weight of this organ, with the inclusion of 30 and 45 % of sugarcane meal replacing alfalfa in diets for rabbits in fattening. Similarly, Albert (2006) observed a similar trend in guinea pigs that received erythrin forage meal, and explained that this effect could be given by the physical-chemical properties of the fibrous source under study and by the reduction of the digesta permanence.
Table 3 Relative weight (g fresh/kg live weight) and percentage contribution of full digestive organs of rabbits fed with moringa forage meal
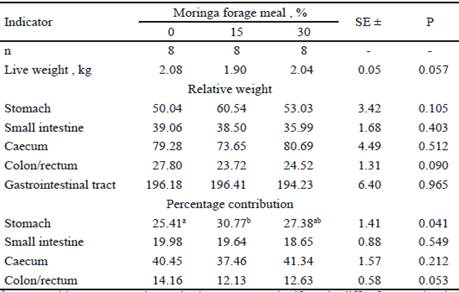
ab Means without common letter in the same row significantly differ from each other (P<0.05)
There was an increase in the percentage contribution of the caecum, as well as a decrease in the relative weight and in the percentage contribution of the empty colon/rectum (table 4) in the animals that intake moringa forage meal with respect to the control. De Blas et al. (2002) suggest that the level and type of dietary fiber influence on the accumulation of digesta in the caecum through its effect on intestinal motility. This performance is determined by the selective process of the fibrous particles that is performed in the caecal-colic segment, as part of the caecotrophy (Gidenne 1996).
Table 4 Relative weight (g fresh/kg live weight) and percentage contribution of empty digestive organs of rabbits fed with moringa forage meal
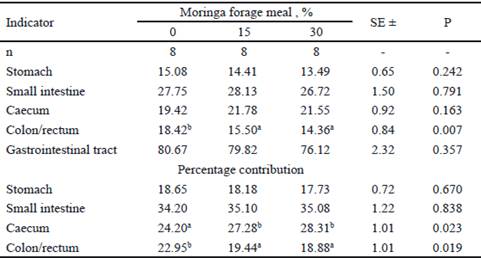
ab Means without common letter in the same row significantly differ from each other (P<0.05)
Table 5 shows the longitudinal measurements of the different segments of the intestinal tract.
Table 5 Longitudinal measurements of empty digestive organs of rabbits fed with moringa forage meal

ab Means without common letter in the same row significantly differ from each other (P<0.05)
The relative length of the small intestine did not differ (P> 0.05) between the treatments in which the different levels of moringa forage meal were included, but with 15 % of this forage differences were observed (P = 0.015) with respect to the control, in which the moringa was absent. In the moringa treatments, the linear density of colon/rectum was lower (P = 0.008). This could be due to the physical- chemical properties of the moringa forage meal. Yu and Chiou (1997) noted that the thickness of the muscle tunic increases because of the distension that causes high levels of fiber in the intestinal segments. In the particular case of moringa, a higher digestibility of the diet (Caro et al. 2018) could imply the presence of lower content of digesta in the posterior regions of the tract, modifying the gastrointestinal architecture as a whole.
Table 6 shows the results corresponding to the fresh digesta of rabbits.
With respect to the relative weight of digesta, this was significantly higher (P = 0.069) in the treatment with 15 % of moringa forage with respect to the control, while these two did not differ from that corresponding to 30 % of tree forage. The percentage contribution of the relative weight of the stomach content showed the same effect (P = 0.025). In contrast, the percent of the relative weight of caecal digesta expressed an opposite effect, with a lower value for 15 % of moringa forage in relation to the treatment without moringa (P = 0.084). This could reflect a change in the site of the gastrointestinal macroarchitecture where the digestive processes of rabbits fed with moringa forage meal take place, with greater importance for the gastric area with respect to the caecal. Dihigo (2007) observed a similar performance when evaluating the effect of different percentages of dehydrated citrus pulp and sugarcane meal on the morphophysiological indicators of the rabbit.
Table 6 Relative weight (fresh g/kg live weight)1 of the digesta content in the digestive organs of rabbits fed with moringa forage meal

ab Means without common letter in the same row significantly differ from each other (P<0.05)
In this study, there was not significant interdependence (R2, 0.004, P> 0.05) between the relative weight of the fresh digesta and the empty segments of the entire digestive tract. Probably this effect was not significant (P> 0.05), due to the high variability found in this measure. There was also no significant effect of the treatment on the relative weight of all fresh digesta (average, 116.7 g/kg live weight). On the other hand, the relative weight of this tract seemed to slightly decrease in a linear way (R2, 0.884, P <0.040) with the introduction of moringa forage in the diet (figure 1).

Figure 1 Interdependence between moringa forage in diet and the digestive tract of rabbits in fattening (Syx, ± 6.46¸P<0.040).
Knowing the gastrointestinal macroarchitecture of the rabbit could have immediate practical consequences, from the point of view not so much of the nutritional physiology as of the carcass yield of rabbits. In this sense, the data provided would be a contribution to this knowledge. It is known that in the rabbit there is an inverse relation between the carcass yield and the weight of the digestive organs (De Blas 1992, Lambertini et al. 2006 and Cornejo et al. 2016). As a demonstration, Kuzelov and Atanosoca (2011) found that the weight of the digestive tract could constitute 17.6 % of the animal weight. In the particular case of rabbits fed with moringa foliage, Dougnon et al. (2012) found that the digestive tract ponderally decreased with the increase of foliage in the food, while the opposite occurred with the carcass yield.
Although the evaluation of the carcasses was not an objective of this study, the data of the gastrointestinal macroarchitecture, from the point of view of the weight of the full digestive organs, could suggest, like those of Dougnon et al. (2012) advantages for a higher yield in the animals carcass. However, Nuhu (2010) found that while the carcass yield was 62 and 66 %, with 0 and 20 % of moringa leaves meal, these two treatments determined 16.4 and 16.3 g/kg of live weight for the weight of the full digestive tract. Due to the limited availability of results on this subject, it is obvious that more research is needed.
It is concluded that the use of moringa forage meal to fatten rabbits can determine few changes in the gastrointestinal macroarchitecture of animals. In this species, the digestive physiology of the use of tree forage from tropical nature, locally available, such as moringa, should be researched in detail.

