










 Custom services
Custom services Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
Meloidogyne spp. es el más dañino de todos los géneros de fitonematodos que atacan al tomate en las regiones tropicales y subtropicales y ocasiona pérdidas en el rendimiento estimadas de hasta un 30 % (Netscher y Sikora, 1990). En Cuba este nematodo, formador de nódulos (rkn), es además uno de los principales problemas fitosanitarios observados en los cultivos protegidos (Gómez, Rodríguez, Enrique, Miranda y González, 2009). Los compuestos químicos pueden controlar esta plaga más rápidamente y con mayor efectividad (Whitehead, 1997). Sin embargo, los daños causados al ecosistema, así como los efectos residuales han traído consigo un aumento del interés para hallar sustitutos más seguros.
En ese sentido, se promueve la aplicación de métodos novedosos y más selectivos para controlar Meloidogyne sp. La búsqueda de antagonistas microbianos naturales puede ofrecer nuevas alternativas para el control biológico de los nematodos. En Cuba varias especies microbianas, como Pochonia chlamydosporia (Manzanilla-López y Lopez-Llorca, 2017) y Tsukamurella paurometabola (Mena et al., 2003), que se utilizan en diferentes cultivos han mostrado una gran eficacia para controlar este tipo de fitopatógeno bajo ciertas condiciones de manejo y tipos de suelo. Otros aislados (Stenotrophomonas sp. CIGB G1 y Sphingobacterium sp. CIGBTb) han demostrado alguna actividad contra zoonematodos en ensayos in vitro (Sánchez et al., 2003) y han mostrado potencial como biocontroles (Sánchez et al., 2018).
El género Sphingobacterium tiene una particularidad llamativa debido a su versatilidad ante diferentes condiciones ambientales. Se ha aislado: del suelo antártico, muestras clínicas, raíces de maíz, heces de ganado vacuno, suelos boscosos y cultivables, lodo activado, nematodos y tejidos de hojas de Nicotiana tabacum (Yabuuchi, Kaneko, Yano, Moss y Miyoshi, 1983; Shivaji et al., 1992; Holmes, Owen y Hollis, 1982; Kim, Ten, Liu, Im y Lee, 2006; Mehnaz, Weselowski y Lazarovits, 2007; Zhang et al., 2012; Liu et al., 2012). Aunque ha sido aislado de suelos con efectos represivos sobre los nematodos, existen pocos reportes sobre su aplicación para controlar el patógeno en tomate.
En concordancia, el objetivo de este estudio fue caracterizar la bacteria CIGBTb aislada de huevos de Tricostrongylus con relación a sus propiedades biocontroladoras sobre los nematodos y sus posibles mecanismos de acción en pruebas en macetas e in vitro, así como la detección de sus propiedades patogénicas.
MATERIALES Y MÉTODOS
Bacterias
CIGBTb se aisló de huevos de Tricostrongylus spp. con una morfología alterada los cuales se desinfectaron con hibitane 0,5 % y fueron colocados en medio agar soya triptona (TSA). El aislado se crioconservó a -70 ºC en la colección del Centro de Ingeniería Genética y Biotecnología de Camagüey, Cuba y su identificación se realizó con el sistema API 20NE y análisis de la secuencia del gen ARNr 16S (números de acceso GenBank: MG461604) obtenida por amplificación de PCR, mediante los cebadores universales 27 F y 1492R (Sánchez et al., 2018). El análisis filogenético se realizó con el empleo del paquete informático MEGA, versión 6,06, después de alineamientos múltiples de datos con CLUSTAL - X (Thompson et al., 1997). Las distancias evolutivas de la cepa CIGBTb se calcularon según el modelo de dos parámetros Kimura (Kimura, 1980), el agrupamiento se basó en el método de máxima probabilidad (Felsenstein, 1981). El análisis bootstrap (1 000 repeticiones) se empleó para evaluar la tipología del árbol (Felsenstein, 1985). La cepa para utilizar en los experimentos se creció en Caldo Soya Triptona (TSB) (Oxoid; 30 g l−1) a 30 ºC, durante 24 h, a 250 rpm, en una zaranda orbital.
El control positivo fue el nematicida Hebernem®, obtenido a partir de la bacteria C924 (depositado el 8 de agosto de 1995, según el Tratado de Budapest, en el Centraalbureau voor Schimmelcultures, Baarn, Holanda, con el número de depósito CBS 613,95.
Los huevos del nematodo Haemonchus sp. se colectaron de hembras adultas presentes en el abomaso de una oveja.
Los especímenes se lavaron con una solución de cloruro de sodio (0,9 %) y se desinfectaron con hibitane al 0,5 % durante un minuto. Luego se incubaron en caldo nutriente (Oxoid), a 37 °C durante 24 horas. Las muestras se tamizaron (300, 60 y 30 µm de diámetro, de manera consecutiva) y los huevos fueron retenidos en el tamiz de 30 µm. Luego, los huevos se desinfectaron con hibitane (0,5 %) durante 5 minutos e inmediatamente lavados dos veces en medio LB (8), diluido 1/10 con agua estéril destilada (LBD). La manipulación se llevó a cabo bajo condiciones asépticas. Los conteos de huevos y larvas se realizaron con un microscopio óptico (Olympus).
Meloidogyne spp. se recolectó de plantas de tomate en cultivos protegidos de la provincia de Ciego de Ávila, Cuba. La población de Meloidogyne spp. se propagó en plantas de Solanum lycopersicum, variedad UC-8 213, en el CIGB de Camagüey, Cuba. Las masas de huevos se recolectaron de las raíces de S. lycopersicum. Los huevos fueron separados de las masas con hipoclorito de sodio (0,5%), y se sumergieron en agua destilada a 80C, hasta la realización del ensayo. Luego se contaron a través de un microscopio binocular invertido (x40) Olympus CK 2. EL conteo de huevos por masa se realizó de la misma manera, pero en cada tratamiento se colocó una masa de huevos en tres pocillos de una placa de poliestireno. Luego los huevos se separaron y finalmente se contaron.
Ensayos en macetas
Se llenaron bolsas de nailon (8 cm de diámetro x 15 cm de profundidad) con 1 000 cm3 de sustrato (1:1 de arena estéril: sustrato estéril enriquecido (Terraplant). El sustrato se infestó con 1 500 huevos de Meloidogyne spp. colocados dentro de las bolsas, a 3 cm de profundidad.
Después de cinco días, se añadió 50 mL de cultivo de Sphingobacterium CIGBTb (106 ufc/mL). El control positivo que se utilizó fue el nematicida Hebernem®, mientras que se utilizó el medio TSB como control negativo. Se empleó un diseño experimental completamente aleatorio, con diez réplicas para cada tratamiento. Las plantas de Solanum lycopersicum UC-8213 fueron trasplantadas a las macetas a los siete días. A los 40 días se determinó el índice de agallamiento, el número de nódulos por gramos de raíz, la altura de la planta, el peso de la raíz y la planta y el número de huevos por masa (Bridge y Page, 1980)
Bioensayo de inhibición de la eclosión de huevos
A cada uno de los 24 pocillos de una placa Petri se le adicionó aproximadamente de 90 a 100 huevos en 900 μL de agua y peptona, a 0,1 % (Meloidogyne spp.) en medio LB diluido 1/10 en agua (Haemonchus). Luego se adicionaron 100 μL de bacterias en un medio de agua/TBS y se probaron tres concentraciones de Sphingobacterium sp. CIGBTb (108, 107, 106 ufc/mL). El control positivo fue Hebernem® en la misma concentración. El control negativo recibió 100 μL de una solución de agua/TBS. Se cubrió la placa y luego se incubó a 28 ºC. El efecto nematicida se determinó con un microscopio binocular, por conteo de los J2s que no eclosionaron a las 72 h del tratamiento. Todos los tratamientos tuvieron tres réplicas. El por ciento de inhibición se calculó según la fórmula PIH = [(C − T)/C] x 100, donde C es el por ciento de eclosión del control y T es el por ciento de eclosión del tratamiento.
Determinación de las enzimas extracelulares
El crecimiento en medio M9 suplementado con quitina coloidal (Shimahara y Takiguchi, 1988) indicó producción de quitinasa. Las quitosanasas se determinaron por el crecimiento del organismo en las placas, con Agar de Detección de Quitosanasa (ADQ) (Cheng y Li, 2000). Las proteasas se determinaron mediante cultivos en las placas con Agar Nutriente y 0,5 % de gelatina, seguido de detección con el reactivo de Frazier y la hidrólisis de caseína en medio agar nutriente con leche descremada al 1 % (VanDemark & Lee, 1991). El sistema Apizym fue empleado para la detección de fosfatasas, esterasas, lipasas, glucoronidasas, galactosidasa, manosidasa, arilasas, glucosaminidasa-β-N-acetil, y fosfohidrolasa AS-BI Naftol.
Los microorganismos se inocularon en tubos que contenían 8 mL de caldo nutriente y se crecieron durante 12 h a 30 ºC. Se colocó una tirilla embebida en acetato de plomo en la apertura del tubo (24). La reacción fue positiva cuando la tira se tornó oscura.
Análisis de datos
Se confirmó la normalidad de los datos con la prueba de Kolmogorov-Smirnov. Se realizó un análisis de varianza (Rodríguez et al. (2005) a todos los datos ensayados para determinar las diferencias significativas entre los tratamientos. Se compararon las medias según la prueba de Duncan (P <0,05). Se empleó el software Statgraphics Plus, 5.0, para Windows.
RESULTADOS
CIGBTb mostró colonias amarillas, convexas y circulares después de dos días de incubación. Las células fueron Gram-negativas a la tinción, en forma de bacilos cortos, inmóviles y no formadores de esporas. El sistema API 20 NE identificó la cepa CIGBTb como Sphingobacterium spiritivorum. El análisis filogenético con las secuencias 16S de ARNr confirmó que este pertenece al género Sphingobacterium. El árbol filogenético de máxima probabilidad mostró que CIGBTb formó un grupo coherente con los miembros del género Sphingobacterium, así como una rama intragénero con Sphingobacterium spriritivorum NCTC 11386 (Fig. 1). Sin embargo, la cepa exhibió un valor de secuencia génica menor de 97 % (95,58 %), por lo tanto, no pertenece a esta especie.

Fig. 1 Árbol filogenético según el método de máxima probabilidad basado en la secuencia de ARNr 16S. El árbol muestra la relación entre la cepa CIGBTb y algunos miembros representativos de la familia Sphingobacteriaceae. Los valores bootstrap (expresados en por cientos de 1 000 repeticiones) por encima de 70 % aparecen en los nódulos de las ramas. La barra representa 2 sustituciones por cada 100 nucleótidos.
El tratamiento in vitro con el empleo de Sphingobacterium sp. CIGBTb, a concentraciones de 108 ufc/mL provocó una inhibición de 87 % en la eclosión de los huevos de Meloidogyne sp. y 100 % de inhibición en la eclosión de los huevos de Haemonchus sp. (Fig. 2). Se observó una gran vacuolización y poca movilidad en las larvas que lograron emerger.
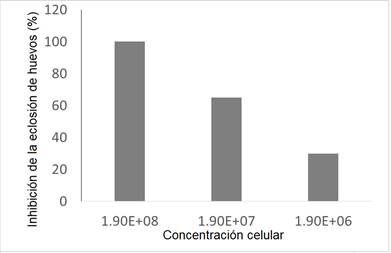
Fig 2 Inhibición en la eclosión de huevos de Meloidogyne spp (A) y Haemonchus spp (B) a diferentes concentraciones de Sphingobacterium CIGBTb
En las macetas, Sphingobacterium sp. CIGBTb redujo significativamente (P < 0,05) el índice de formación de agallas, de 5,3 a 2,9. No se observaron diferencias significativas en el control positivo. Por otra parte, Sphingobacterium sp. CIGBTb disminuyó significativamente las formaciones de agallas (58 %) en las raíces de Solanum lycopersicum (Tabla 1).
Tabla 1 Efecto de Sphingobacterium sp. CIGBTb en las raíces de S. lycopersicum UC-8213 infestado con Meloidogyne spp.
| Tratamiento | Severidad de los nódulos | ||
|---|---|---|---|
| Índice de formación de nódulos | Nódulos/g de raíz | ||
| CIGBTb | 2,9 ± 1,5 b | 27,8 ± 4,8 b | |
| Control | 5,3± 1,2 a | 47,8 ± 2,8 b | |
Letras diferentes representan diferencias significativas en la prueba de Duncan (P < 0, 05).
Una evaluación del número de huevos por masa, aislados de los diminutos nódulos que se formaron, mostró que Sphingobacterium sp. CIGBTb redujo el número de huevos en cada masa (53 %), mientras que no se observaron cambios en los controles (Fig. 3).
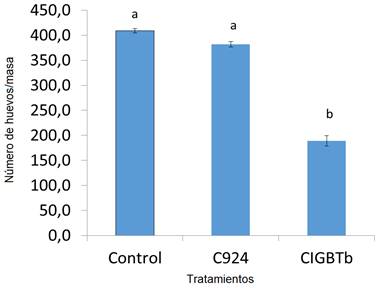
Fig 3 r Número de huevos por masa en las plantas tratadas con caldo de soya triptona (control) Sphingobacterium sp. CIGBTb (CIGBTb) y Hebernem® (C924) Letras diferentes epresentan diferencias significativas en la prueba de Duncan (P < 0, 05).
Una concentración de aproximadamente 105 ufc/mL, Sphingobacterium sp. CIGBTb también estimuló el crecimiento de Solanum lycopersicum. También aumentó el peso de la planta en 0,59 veces por encima del peso de las plantas del tratamiento control sin la bacteria (Fig. 4).
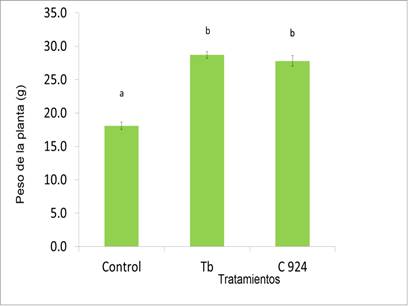
Fig 4 Peso de plantas de S. lycopersicum UC-8213 infestadas con Meloidogyne spp., 40 días después del tratamiento con caldo soya triptona (control), Sphingobacterium sp. CIGBTb (CIGBTb) y Hebernem® (C924) Letras diferentes representan diferencias significativas en la prueba de Duncan (P < 0, 05).
El CIGBTb presentó varios posibles atributos de patogenicidad, como tripsina, esterasas y enzimas lipasa estereasas. El sulfuro de hidrógeno y las proteasas extracelulares (con la gelatina y la caseína como sustratos) no fueron detectados mediante la prueba convencional y la cepa logró crecer lentamente en medio de cultivo mínimo con quitina y quitosano, aunque no se observó la formación de halo de hidrólisis. Además, los resultados ofrecidos por el sistema Apizym fueron positivos para la fosfatasa ácida y la fosfatasa alcalina, dos enzimas importantes para la solubilización de fósforo en el suelo por medio de las bacterias.
DISCUSIÓN
Anteriormente se había reportado el género Sphingobacterium como un componente de la rizosfera de vid con propiedades supresoras de nematodos (Vargas-Ayala, Rodrı́guez-Kábana, Morgan-Jones, McInroy, & Kloepper, 2000; Aballay, Mårtensson, & Persson, 2011). Estos últimos demostraron la efectividad de la cepa de Sphingobacterium spiritivorum 64 in vitro, así como otra cepa del género (Sphingobacterium nematocida) que se aisló en China durante un estudio sobre la diversidad de los organismos nematicidas endofíticos (Liu et al., 2012). No obstante, existe poca información sobre los ensayos en maceta y los posibles mecanismos de acción de estos géneros de bacterias. Mena et al. (1996) patentó la cepa de Sphingobacterium spiritivorum C926 y demostró su efectividad para controlar Radopholus similis y Meloidogyne incognita en ensayos de campo y en macetas. Sin embargo, Sphingobacterium CIGBRTb no solo tiene actividad nematicida, sino que también estimula el crecimiento de plantas de tomate, posiblemente debido a la solubilización de los fosfatos del suelo, mediante la producción de fosfatasas ácidas y alcalinas, aunque otros mecanismos no han sido estudiados. Anteriormente se conocía la actividad estimuladora del crecimiento de las plantas por parte de Sphingobacterium canadense (Mehnaz et al., 2007) y Sphingobacterium pakistanense (Ahmed et al., 2014).
Existen varios mecanismos que utilizan las bacterias durante interacciones antagonistas con los nematodos. Uno de ellos es el papel que desempeñan las enzimas hidrolíticas, lo cual está bien documentado para una serie de organismos con actividad híperparásita (Chernin & Chet, 2002). En el caso de Sphingobacterium CIGBTb, la cepa tenía actividad quitinasa, quitosanasa, tripsina, esterasa y lipasa esterasa. Estos atributos patogénicos pudieran traer consigo cambios enzimáticos en las capas de quitina, las proteínas y otros lípidos que conforman la cubierta de los huevos y otras estructuras de Meloidogyne sp., y por consiguiente, facilitan la ocurrencia de parasitismo.
Existen varias cepas de Sphingobacterium que producen quitosanasas similares a Mitsuaria ChoA (Yun, Amakata, Matsuo, Matsuda, & Kawamukai, 2005) y esta cepa en particular pudiera ser parte de ellas. Además, CIGBTb libera esterasas C4 y C8, como el Sphingobacterium nematocida (Liu et al., 2012), que pudieran explicar la inhibición de la eclosión in vitro de huevos de Meloidogyne sp. Sin embargo, las larvas que logran emerger y parasitar las plantas de tomate en los ensayos de maceta no solo producen un bajo número de agallas, sino también que estas son más pequeñas, con menos huevos en las masas que el control. El silenciamiento de un gen de parasitismo, el 16D10, expresado en las células de las glándulas subventrales de M. incognita también provoca un efecto similar en Arabidopsis. Se produce una reducción sustancial del número de agallas entre 63 y 90 %, así como en el tamaño de las agallas (Huang, Allen, Davis, Baum y Hussey, 2006).
CONCLUSIONES
Los zoonematodos son una fuente apropiada para la búsqueda de biocontroladores de nematodos fitopatógenos. Sphingobacterium sp. CIGBTb aislado de huevos de Tricostrongylus sp. es una alternativa novedosa para el control biológico. La cepa reduce el número de huevos por masa, a diferencia del ingrediente activo de C924 del producto nematicida Hebernem®, que libera quitinasas y sulfuro de hidrógeno. Los resultados de este estudio corroboraron la efectividad del aislado nativo como antagonista, así como su potencial como biofertilizante de las plantas y control biológico del nematodo Meloidogyne sp., formador de nódulos en Solanum lycopersicum

