










 Custom services
Custom services
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
Juglans jamaicensis C. DC. subsp. jamaicensis (Juglandaceae) es una especie endémica de Las Antillas, con categoría de amenaza en peligro, según los criterios de los autores González y otros., (2016). Este forma parte del primer estrato arbóreo de los bosques húmedos semicaducifolios en lugares de altas precipitaciones y buena retención de agua, preferentemente de suelos arcillosos bien drenados, aunque se han encontrado en suelos de características variadas, incluso, ligeramente alcalinos según explica Betancourt, (1999). Este mismo autor indica que la especie habita naturalmente entre los 200 y 300 metros de altitud hasta los 1 100 m. de altitud en La Española.
Varios autores como Betancourt, (1999); Bibb y Monsegur, (2013) refieren declive de sus poblaciones, así como alteración de su hábitat por el cultivo del café, extracción irracional por la calidad de su madera, a lo cual se suman los aludes o deslizamientos de suelo en las montañas y el impacto del cambio climático manifestado a través del incremento de los eventos meteorológicos extremos como son los huracanes.
Aunque en Cuba se ha protegido la subespecie y se han trazado un grupo de estrategias para la conservación de la misma, asentadas en el programa de Desarrollo Forestal hasta el 2015 de la provincia de Granma, aún persisten escasa regeneración e individuos deprimidos por lianas, así como tala ilegal de árboles longevos en condiciones naturales según lo expresado por Rodríguez, (2015). Por lo tanto, el objetivo de este trabajo es caracterizar la distribución espacial de las semillas de la especie en el Parque Nacional Turquino, área protegida, de significación nacional.
Materiales y métodos
El área de estudio comprendió fragmentos remanentes de bosques de montaña afectados por el cultivo del cafeto y su abandono, así como por los deslizamientos de tierra, ubicados en las Unidades Zonales de Conservación Santo Domingo y La Platica, pertenecientes al Parque Nacional Turquino.
La comunidad donde vive la especie se caracteriza por presentar una estructura horizontal determinada, de acuerdo con su mayor peso ecológico, por Dendropanax arboreus (L.) Decne., Prunus occidentalis Sw., Ficus membranacea C. Wright., Erythrina poeppigiana (Walp.) O.F. Cook, Guarea guidonia (L.) Sleumer, Sapium jamaicense Sw., Beilschmiedia pendula (Sw.) Hemsl., Zanthoxylum martinicense (Lam.) DC., Prestoea montana (R. Graham) G. Nicholson, Ocotea leucoxylon (Sw.) Laness., Licaria jamaicensis (Nees) Kosterman y Trema micranthum (L.) Blume. Rodríguez, (2015).
Esta comunidad se favorece con un clima donde las precipitaciones son superiores a los 1 600 mm. y las temperaturas oscilan entre los 21 y 24 oC., así como un suelo ferralítico rojo lixiviado de profundo a muy profundo y ácidos. Reyes, (2011).
La tabla 1 reúne la información geográfica y topográfica de los sitios con árboles de Juglans jamaicensis C. DC. subsp. jamaicensis.
Todos los sitios se ubicaron junto a corrientes pequeñas de agua que favorecen el alto contenido de humedad en el ambiente. Se seleccionaron siete grupos de árboles de J. jamaicensis en las UZC Santo Domingo y La Platica. (Tabla 1)
Tabla 1 - Ubicación geográfica de los grupos de árboles de Juglans jamaicensis C. DC. subsp. jamaicensis en las Unidades Zonales de Conservación Santo Domingo y La Platica del Parque Nacional Turquino.
| Unidad Zonal de Conservación | Sitio | Número de árboles | Ubicación geográfica | Altitud (msnm) | Pendiente (o) | |
| Latitud N | Longitud W | |||||
| La Platica | PM | 2 | 20o 00´ 13,1´´ | 76o 52´ 44,3´´ | 1266 | 33,0 |
| RA | 1 | 20o 00´ 16,4´´ | 76o 53´ 0,02´´ | 1144 | 22,5 | |
| RB | 1 | 20o 00´ 17,3´´ | 76o 53´ 5,6´´ | 1092 | 35,1 | |
| Santo Domingo | AO | 1 | 20o 00´ 55,4´´ | 76o 52´ 55,9´´ | 952 | 44,8 |
| JA | 1 | 20o 02´ 1,8´´ | 76o 50´ 38,3´´ | 745 | 16,5 | |
| JB | 1 | 20o 02´ 5,2´´ | 76o 50´ 48,3´´ | 595 | 10,8 | |
Diseño de muestreo para la caracterización de la dispersión de semillas de Juglans jamaicensis
En cada árbol, se determinó la dispersión de las semillas en parcelas de 1 x 1 m., ubicadas cada 2 m. a lo largo de cuatro transectos radiales de 50 m. de longitud, orientados hacia los cuatro puntos cardinales, desde la base del árbol donante de semillas. Parrado, (2007).
Las semillas fueron recolectadas de la superficie del suelo de forma manual y posteriormente trasladadas hasta la Estación Experimental Agroforestal Guisa donde fueron puestas a germinar en el germinador, al aire libre, alcanzando como promedio 73 % de capacidad germinativa. Rodríguez, (2015).
Resultados y discusión
La densidad de las semillas de J. jamaicensis dispersadas en la superficie del suelo declinó marcadamente con la distancia desde la planta parental, y se ajustó significativamente su distribución a una ecuación polinómica de segundo orden. (Figura 1)
La mayor densidad de semillas ocurrió en las cercanías de las plantas progenitoras, lo que determinó para J. jamaicensis un alcance máximo de 23 m. en dispersión primaria de semillas (Figura 1), mientras que la distancia media de dispersión, alrededor de los árboles, fue de 13 m. y albergó el 84 % de las semillas.
Las distancias estimadas fueron superiores a las encontradas para distintas especies arbóreas en otros estudios procedentes de bosques tropicales donde habitan especies del género Juglans. Así, el alcance máximo en la dispersión primaria de semillas de Pseudolmedia oxyphyllaria fue de 25 m., según lo expresado por los autores Mostacedo y Pinard (2001); 7,1 m. en Dacryodes chimantensis y 12 m. en Brosimum utile.Parrado, (2007); inferiores a los 125 m. en Spondias radlkofirii . Mostacedo y Pinard, (2001); en Juglans cinerea L., el máximo es de 168 m. como consecuencia de la acción dispersora de las ardillas. Tamura y otros., (1999).

La corta distancia de dispersión de J. jamaicensis ocasiona un restringido movimiento en la población que podría explicar el patrón agregado encontrado para la especie en el Parque Nacional Turquino por Rodríguez, (2015), similar también al reportado por Wang y otros., (2008) para Juglans mandshurica Maxim., como consecuencia de la lluvia de semillas, alrededor de los árboles progenitores.
En efecto, esta disposición de las semillas desde el punto de vista espacial podría generar un reclutamiento bajo, ya que según Dalling, (2002) este comportamiento reduce, en gran medida, las posibilidades de que la especie domine a nivel local y ceda el espacio a otras para que recluten sus plántulas y, por otra parte, también posibilitaría la depredación de semillas en las cercanías del árbol progenitor como han reportado en otros estudios Terborgh y otros., (1993).
La dispersión restringida de semillas acarrea consecuencias genéticas importantes, porque el establecimiento local de propágulos resulta en un agrupamiento de individuos genéticamente emparentados en la cercanía del árbol semillero, y que, en algunos casos, ocurre a escalas espaciales reducidas (< 20 m), como en los bosques maduros de Nothofagus dombeyiRovere y Premoli, (2005).
En este caso, y a partir de evidencias de actividad química alelopática de varias especies del género JuglansWillis, (2000), Hassannejad y Ghafarbi, (2013); Khoshvaghti y Lotfi, (2013), podrían sugerir que la dispersión de semillas tan contagiosa, bajo la planta madre, estuviese interfiriendo químicamente entre el árbol adulto y las plantas juveniles, ya que Hepting, (1971) y Schultz, (2003) expusieron que al igual que otras especies del género, Juglans cinerea L., exhibe tendencias alelopáticas, al exudar de sus raíces una sustancia llamada juglona que es tóxica a muchos otros árboles e incluso a sus propios arbolillos.
Por otra parte, la dispersión de semillas de J. jamaicensis varió entre los puntos cardinales, lo que generó un patrón de distribución asimétrico con mayor tendencia hacia el norte, como consecuencia de la pendiente del terreno, lo que evidencia la intensa influencia que sobre la sombra de semillas pueden ejercer las características orográficas locales.
Esta asimetría de la sombra de semillas puede asociarse, además, a la topografía, la heterogeneidad del hábitat Marone y otros., (2004) u otros factores ecológicos, como puede ser el patrón de comportamiento de vectores dispersores de semillas. Tackenberg y otros., (2003); Rovere y Primoli, (2005). (Figura 2)
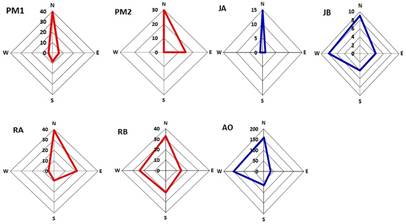
Fig. 2 - Dispersión de las semillas de los árboles progenitores en relación con los puntos cardinales
Si bien la distancia de dispersión para J. jamaicensis resultó aproximadamente de 23 m., consideramos que la distancia real de máxima dispersión puede ser superior, ya que el método de muestreo usado no consideró potenciales eventos de dispersión secundaria de las semillas, o sea, desplazamientos desde un punto de partida distinto del árbol progenitor, típicamente el suelo, causado en muchos casos por roedores como exponen García, (2002) en Juglans olanchana Standl. & L. O. Williams.,Schultz, (2003) para Juglans cinerea L.,Tamura y Hayashi, (2008) para Juglans ailanthifolia Carr.; Blendinger y Díaz, (2010) en Juglans australis Griseb; Yi y Yang, (2011) en semillas de Juglans mandshurica Maxim., y Bibb y Monsegur, (2013) para J. jamaicensis en Puerto Rico. En este último caso, al encontrar frutos sin pericarpio y semillas bajo las rocas, al igual que en los sitios Altos de Palma Mocha y Rolando (Figura 3A), sugieren la intervención de animales en la dispersión de las semillas, como por ejemplo la rata negra, múrido introducido y superabundante en todas las áreas boscosas de Cuba Borroto y Mancina, (2011), que genera este tipo de patrones espaciales de dispersión secundaria.
Esta rata también constituye, probablemente, el consumidor de un gran número de semillas bajo las rocas cercanas a los árboles, pues Janzen y Vázquez, en el 1991, citados por Dalling, (2002), refieren que cerca de la mitad de las semillas producidas por más del 90 % de todas las especies de árboles del bosque tropical, mueren antes de germinar, presas de animales y hongos.
Otro de los mecanismos que tributa a la dispersión de las semillas en la Sierra Maestra, es el agua. (Figura 3B); al encontrarse los árboles generalmente junto a corrientes de agua, en la montaña. Al respecto, reportó Schultz, (2003) que existen las siguientes variedades de los árboles: J. cinerea en las fajas de bosque junto a los ríos del centro-este de Estados Unidos, y al sureste de Canadá, y J. jamaicensis en la Sierra Maestra.
Esta tendencia puede fundamentarse también por los mecanismos de dispersión de la especie, reportados por Schaarschmidt, (2002) y Bibb y Monsegur, (2013) como bolocora, hidrocora, zoocora y antropocora.
Teniendo en cuenta lo anterior y conociendo que la autocoría es un mecanismo de dispersión únicamente relacionado con el árbol progenitor, que deja caer las semillas maduras Noir y otros., (2002), es importante reconocer que las especies autocóricas probablemente dependen de un dispersor secundario, pues según Van der Pijl en el año 1982, citado por Matsumoto, (2009) Miranda y otros., (2005), muchas no disponen de mecanismos eficientes para la dispersión.

Fig. 3 - Acumulación de semillas bajo rocas (A) y en el curso de agua (B) como indicador de los factores bióticos y abióticos que pueden influir en la dispersión de las semillas de Juglans jamaicensis en el Parque Nacional Turquino
La distribución de las semillas en la superficie del suelo se comportó a razón de 12 semillas por m2 (banco de semillas pequeño (<30 sem/m2), Zhang y otros., (1997), inferior a la obtenida por Ferrandis y otros., (2011) para la especie amenazada Podocarpus angustifolius Griseb (283 semillas/m2). Este resultado sugiere un grado sustancial de fragilidad de la especie frente a perturbaciones y factores externos, aunque Siqueira (2002) refiere que el número limitado de semillas de la mayoría de las especies tropicales se debe principalmente a la baja longevidad y ausencia de dormancia que permanecen poco tiempo disponible en el banco.
Siendo así, el conocimiento de la dinámica y densidad del banco de semillas ofrece indicaciones sobre la resiliencia de determinada área y permite anticipar el efecto de prácticas de manejo y de los disturbios naturales o antrópicos, Onaindia y Amezaga, (2000), así como facilitar, por medio de la regeneración natural, la recomposición de la cobertura vegetal en áreas degradadas. Costalonga, (2006).
Juglans jamaicensis mostró un patrón de dispersión contagioso que declina marcadamente con la distancia desde la planta parental.
La sombra de semillas asimétrica estuvo asociada a la influencia de las características orográficas locales y al comportamiento de vectores dispersores de semillas como el agua de los arroyos y los animales.
El patrón espacial en la dispersión primaria de las semillas de Juglans jamaicensis, así como su baja densidad, causante de la formación de un banco pequeño de semillas, constituyen elementos indicadores de la vulnerabilidad de las poblaciones frente a posibles perturbaciones, así como del patrón de distribución agregado de la especie en el bosque.

