










My SciELO
 Custom services
Custom servicesServices on Demand
Article
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista Cubana de Ciencias Forestales
On-line version ISSN 2310-3469
Rev cubana ciencias forestales vol.9 no.1 Pinar del Río Jan.-Apr. 2021 Epub Apr 07, 2021
Original article
Resource use by lizard communities in semi-deciduous forests and its relationship with vegetation variables under different harvesting times
1Universidad de Pinar del Río "Hermanos Saíz Montes de Oca". Pinar del Río, Cuba.
2Centro de Investigación y Servicios Ambientales. ECOVIDA. Pinar del Río, Cuba.
Reptiles occupy a great variety of habitats and types of ecomorphs, and are among the taxonomic groups most sensitive to disturbances, such as habitat fragmentation and loss. The objective of this study was to evaluate the incidence of changes in the semi-deciduous forest, in the dynamics of recovery after forest harvesting interventions, on the use of certain resources in lizard communities and their relationship with vegetation variables. An observation guide was used in the semi-deciduous forest in two treatments with different post-logging recovery times in two sectors of the Guanahacabibes National Park, Cabo San Antonio and Cabo Corrientes, to determine the lizard species, the illumination to which they are exposed, the type of substrate and the height frequented by each individual. Thirty-six species were identified; the use of resources by the detected species varied significantly with post-harvest recovery time for height and illumination. The vegetation variables that showed the greatest association with the different species of lizards were: species richness, density of the lower tree stratum and height of the three established strata.the associations established between changes in forest structure and lizard communities are significantly manifested by the strata of height and illumination in both locations and the vegetation variables that have a greater correspondence are richness of the shrub stratum, height of the upper tree stratum and natural regeneration.
Key words: Forest use; Semi-deciduous forest; Ecomorphs and resource.
INTRODUCTION
Within vertebrates, reptiles are among the taxonomic groups most sensitive to disturbances, such as habitat fragmentation and loss and the local and global effects of climate change, especially in insular groups (Allison et al., 2002). Cuban terrestrial reptiles are represented by 160 species, of which 64 belong to the genus Anolis (Losos 2009 and Rodríguez Schettino et al., 2013).
Reptiles occupy a wide variety of habitats and cover types, from open areas and transformed forests to preserved forests (Duellman 1979); this differential use of the original habitats or covers gives rise to different horizontal and vertical distribution patterns of the assemblages, with the consequent differentiation in the use of resources, including food (Pianka 1989; Duellman 1979; Vitt and Pianka 2005; Mesquita et al., 2006).
The composition of the lizard fauna is profoundly influenced by vegetation structure. In general, the habitat requirements of almost all reptile species in the Antilles are defined by morphological, physiological and behavioral adaptations (Schwartz and Henderson 1991).
When changes occur in the landscape, due to anthropogenic actions such as fragmentation and loss of vegetation cover, the availability, abundance and richness of the available resource are affected (Carvajal and Urbina-Cardona 2008). This situation has repercussions on the response and adaptation of species to the new conditions of use of the available resources.
Based on these theoretical facts, this research aims to evaluate the incidence of semi-deciduous forest modifications on the dynamics of recovery after forest harvesting interventions, on the use of certain resources in lizard communities and their relationship with vegetation variables.
MATERIALS AND METHODS
The research was carried out in the semi-deciduous forest in two sectors of the Guanahacabibes National Park: Cabo San Antonio and Cabo Corrientes, located at the westernmost extreme of the island of Cuba (Figure 1).
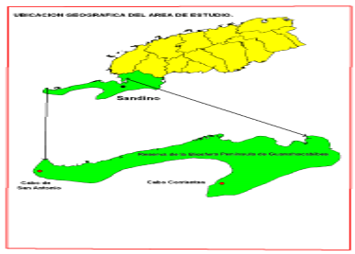
Fig. 1 - Location of the study localities, Cabo de San Antonio and Cabo Corrientes in the Guanahacabibes Peninsula
To evaluate the impacts of forest modifications on lizard communities. Two treatments were evaluated in each sector with different post-logging recovery times during three years. (2013, 2014 and 2015):
Cabo de San Antonio (CSA) and Cabo Corrientes (CC).
Treatment 1 - 5 years of post-logging recovery.
Treatment 2 - 30 years or more of post-logging recovery.
In each treatment, 10 transects with a length of 100 m and a width of 5 m were made, at a distance of 100 m from each other following the method of Rand (1964) . They were walked at a slow pace from 9:00 to 13:00 hours in two periods of the year, rainy and not very rainy, and the following data were collected:
Sex and stage (adult or juvenile females and males).
Type of substrate where the individual is found when observed, tree or shrub trunk, rock, branch or soil.
When the substrate is a branch or trunk, the height from the ground is measured with a tape measure graduated in 0.5 cm and the diameter of the part that supports it.
Illumination to which the animal is exposed, classifying it in filtered sun (F), direct sun (D) and shade (S).
Data analysis
A study of the strength of the relationship between the potential predictor variables and the dependent variable was carried out using Spearman's rank correlation coefficient. Among multiple models tested, it was decided to select the model that includes as intrasubject variables, locality, season, treatment and year.
Binary logistic regression was used for the associations between the variables (perch height, illumination and substrate frequented).
The Mantel-Haenszel statistic was applied in the multivariate analysis to estimate the common advantage controlling for the strata of height, illumination and substrate. The Chi-square statistic (c2) was also used to verify the significance of the homogeneity of the advantage ratio between the controlled strata using the Breslow-Day and De Tarone tests.
By means of the canonical correspondence analysis, using the program PcOrd version 4.10, the associations established between the densities of the lizards and the measured variables of the forest structure are analyzed in a direct way.
The Mantel test, which is widely used in the analysis of multivariate ecological data, is performed. It is a regression in which the variable matrices are distance or similarity matrices, which summarizes the similarities in many variables between pairs of localities. This statistic, however, does not have a fixed theoretical distribution but is compared to the null distribution obtained by permutations of rows and columns of the data matrix (null model).
RESULTS AND DISCUSSION
Effect of post-harvest recovery on the use of resources
Perch height
Table 1 shows the associations between the categorical variables weighted by the frequencies of the variables stated in the first column. The variables with which the associations with the dependent variable were calculated are presented, using binary logistic regression. The odds ratio and its confidence interval are also presented and the last column of the table shows the associations that were significant, as well as the bilateral significance.
Table 1 - Degree of association between categorical variables. Total lizards
| Weighting variable | Associations | Reason for advantages | Lower limit | Upper Limit | P |
| frequency by height | location CC/CSA | ||||
| Alt_1.99/Alt_0.99 | 0.836 | 0.714 | 0.979 | 0.026* | |
| Alt_2.00/ Alt_0.99 | 0.714 | 0.979 | 0.714 | 0.027* | |
| Año 2007/2006 | 0.770 | 0.645 | 0.919 | 0.004* | |
| Año 2008/2006 | 0.764 | 0.643 | 0.908 | 0.002* | |
| treatments | |||||
| Alt_1.99/Alt_0.99 | 1.155 | 0.987 | 1.352 | ns | |
| Alt_2.00/ Alt_0.99 | 1.101 | 0.848 | 1.431 | ns | |
| Año 2007/2006 | 0.946 | 0.792 | 1.129 | ns | |
| Año 2008/2006 | 1.024 | 0.862 | 1.218 | ns | |
| Season | |||||
| Alt_1.99/Alt_0.99 | 1.003 | 0.857 | 1.173 | ns | |
| Alt_2.00/ Alt_0.99 | 0.724 | 0.555 | 0.943 | 0.017* | |
| Year 2007/2006 | 0.770 | 0.645 | 0.919 | 0.004* | |
| Year 2008/2006 | 0.764 | 0.643 | 0.908 | 0.002* | |
| Location*treat. | 1.118 | 0.971 | 1.286 | ns | |
| Treat.*Season | 0.934 | 0.812 | 1.075 | ns | |
| Treat. *years (2007) | 0.946 | 0.792 | 1.129 | ns | |
| Treat. *years (2008) | 1.024 | 0.862 | 1.218 | ns | |
| Season*years (2007) | 0.953 | 0.798 | 1.137 | ns | |
| Season*years (2008) | 0.956 | 0.804 | 1.136 | ns | |
| Frequency by illumination | Location CC/CSA | ||||
| Filtering /sun * | 1.447 | 1.222 | 1.715 | <0.001* | |
| Shadow /sun | 1.080 | 0.894 | 1.304 | ns | |
| Year 2007/2006 | 0.735 | 0.616 | 0.878 | 0.001* | |
| Year 2008/2006 | 0.742 | 0.624 | 0.883 | 0.001* | |
| Treatment | |||||
| Filtering/sun | 0.890 | 0.752 | 1.053 | ns | |
| Shadow/sun | 1.026 | 0.850 | 1.237 | ns | |
| Year 2007/2006 | 0.735 | 0.616 | 0.878 | ns | |
| 2008/2006 | 0.742 | 0.624 | 0.883 | ns | |
| Season | |||||
| Filtering/sun* | 1.203 | 1.016 | 1.424 | 0.032* | |
| Shadow/sun* | 1.232 | 1.021 | 1.487 | 0.029* | |
| Year 2007/2006 | 0.963 | 0.807 | 1.149 | ns | |
| Year 2008/2006 | 0.982 | 0.826 | 1.167 | ns | |
| Location*treat. | 0.902* | 0.783 | 1.038 | ns | |
| Treat.*Season | 1.057 | 0.919 | 1.216 | ns | |
| Treat. *Years (2007) | 0.926 | 0.776 | 1.105 | ns | |
| Treat. *Years (2008) | 1.005 | 0.846 | 1.195 | ns | |
| Season*Years (2007) | 0.963 | 0.807 | 1.149 | ns | |
| Season*Years (2008) | 0.982 | 0.826 | 1.167 | ns | |
| Frecuency by substrate | Location CC/CSA | ||||
| Rock/soil | 1.193 | 0.930 | 1.530 | ns | |
| Trunk/soil | 0.863 | 0.711 | 1.047 | ns | |
| Branch/soil | 0.000 | 0.000 | - | ns | |
| Year 2007/2006 | 0.898 | 0.747 | 1.078 | ns | |
| Year2008/2006 | 0.866 | 0.725 | 1.035 | ns | |
| treatment | |||||
| Rock/soil | 1.036 | 0.808 | 1.327 | ns | |
| Trunk/soil | 1.070 | 0.882 | 1.299 | ns | |
| Branch/soil | 6.982 | 2.689 | 18.129 | <.001* | |
| Year 2007/2006 | 0.873 | 0.727 | 1.049 | ns | |
| Year 2008/2006 | 1.045 | 0.874 | 1.249 | ns | |
| Season | |||||
| Rock/soil | 0.769 | 0.599 | 0.986 | 0.038* | |
| Trunk/soil | 0.775 | 0.639 | 0.941 | 0.010* | |
| Branch/soil | 0.469 | 0.238 | 0.922 | 0.028* | |
| Year 2007/2006 | 1.036 | 0.863 | 1.245 | ns | |
| Year 2008/2006 | 0.963 | 0.806 | 1.152 | ns | |
| Loctlion* treat. | 0.910 | 0.786 | 1.053 | ns | |
| Treat. *Season | 1.123 | 0.970 | 1.300 | ns | |
| Treat. *Years (2007) | 0.873 | 0.727 | 1.049 | ns | |
| Treat. *Years (2008) | 1.045 | 0.874 | 1.249 | ns | |
| Season* Years (2007) | 1.036 | 0.863 | 1.245 | ns | |
| Season* Years (2008) | 0.963 | 0.806 | 1.152 | ns |
The multivariate analysis (Table 2), showed significant differences in the combinations "Location* treatment" controlled by the height strata with a common advantage ratio of 0.9 and a significance in the homogeneity of advantages test of 0.018, which shows that the behavior is not the same in the different strata. The same is true for "Location * season" with an odds ratio of 1.085 which is the number of times that lizards are more likely to be found at one station than another when controlling for height and a significance in the homogeneity of odds test of 0.033.
Table 2 Results of association in multivariate análisis
| Associations | Mantel-Haenszel statistic | Significance of benefit ratio homogeneity | |||
| Common benefitratio | Bilateral asymptotic significance | Statistic (2 | Bilateral asymptotic significance | ||
| Frequency by height | Location*Treatment*Season | 1.094 | 0.213 | 0.043 | 0.836 |
| Location*Treatment* Years | 0.894 | 0.119 | 1.964 | 0.375 | |
| Location*Season* Year | 1.089 | 0.237 | 0.514 | 0.773 | |
| Season* Treatment* Years | 1.070 | 0.363 | 1.316 | 0.518 | |
| LocationTreatment*height | 0.900 | 0.143 | 8.013 | 0.018* | |
| Location*Season* height | 1.085 | 0.254 | 6.80 | 0.033* | |
| Season* Treatment*height | 1.072 | 0.334 | 0.274 | 0.872 | |
| Frecuency by Illumination | Location*Treatment* Season | 0.900 | 0.144 | 0.221 | 0.638 |
| Location*Treatment* Years | 0.899 | 0.139 | 2.083 | 0.149 | |
| Location*Season *Years | 1.087 | 0.248 | 1.251 | 0.263 | |
| Season* Treatment* Years | 1.057 | 0.443 | 1.662 | 0.436 | |
| Location*Treatment*Illumination | 0.910 | 0.192 | 9.837 | 0.007* | |
| Location*Season *Illumination | 1.079 | 0.291 | 1.345 | 0.510 | |
| Season* Treatament*Illumination | 1.059 | 0.422 | 0.079 | 0.961 | |
| Frecuency by substrate | Location*Treatment*Season | 0.908 | 0.196 | 0.390 | 0.532 |
| Location*Treatment* Years | 0.910 | 0.206 | 4.040 | 0.133 | |
| Location*Season* Year | 1.075 | 0.335 | 2.917 | 0.233 | |
| Season* Treatament* Year | 1.126 | .114 | 1.066 | 0.114 | |
| Location*Treatment*Substrate | 0.947 | 0.467 | 0.206 | 0.902 | |
| Location*Season* Substrate | 1.058 | 0.453 | 21.670 | 0.001* | |
| Season* Treatment* Substrate | 1.139 | 0.084 | 2.890 | 0.409 | |
*significant differences at 5%. Source: Own elaboration based on SPSS output tables.
Height
Weighting: frequency by height
The height category of greatest use in all combinations of the 3 effects considered (year, season and treatments) for both localities was 0-0.9 m (Figure 2). With locality the associations in the height and year categories were significant, in the first case the number of times that lizards were likely to be found at the height of 1.99 with respect to 0.99 m was 0.836. They did not indicate significant associations between treatments for any of the year*season combinations. Season showed significant association with the frequency of lizards at heights equal to or greater than 2.00 m and with years. The rest of the associations when weighted by frequency according to height were not significant.
In both locations there was a trend towards a decrease in the use of the 0-0.9 m category. By 2013, from the rainy period onwards, the use of this category established in the 5-year recovery treatment began to decrease slightly, which continued more markedly in 2014. This was influenced by the increase in higher frequency of lizards due to the use of the intermediate height (1.00-1.99 m). This same pattern began in 2014 in the rainy period for the treatment of more than 30 years without forest harvesting. For the year 2015, in both seasons, the differences between uses of height 0-0.9 m were smaller than in previous years.
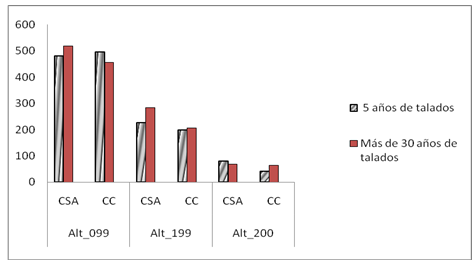
Fig. 2 - Frequency of lizards. Location Cabo de San Antonio (CSA) and Cabo Corrientes (CC) by Treatment by the three established perch height categories.
In both localities, the preferred height for lizards was 0-0.9 m, with a tendency to decrease the use of this category in the two treatments for the last two years, increasing the number of individuals in the second category established (1,00-1,9 m). In the Cabo Corrientes locality, significant differences appear from 2013 during the rainy season and continue throughout 2014 in both seasons, and in the Cabo de San Antonio locality during the dry season in the last year of sampling. This response is due to the fact that the lizards are observed occupying individuals of the natural regeneration of the forest, less than 2 m in height and less than 2 cm in diameter, established by Delgado and Ferro (2000), which coincides with the one preferred by the lizards of the community under study.
This natural regeneration varies depending on the treatment given to the forest, in more disturbed places there is a more significant variation in terms of density in the rainy and less rainy periods, in the treatments with less time of forest harvesting it is defined by the seed bank and in the more conserved treatments the regeneration strategy is by seedlings.
In cases where both anthropic and natural disturbances cause clearings in the forest due to a decrease in vegetation cover, as in the case of the treatment with less time of recovery after forest harvesting, the regeneration behavior is totally different. Forest management causes a decrease in the diversity of forest species in the forest and does not activate the increase of them in the natural regeneration (Delgado and Ferro, 2000).
From a forestry point of view, at higher altitudes of the strata there is a notable increase in the tree canopy, bringing with it a greater density of the canopy and with it a decrease in sunlight penetration (Delgado and Ferro, 2006). Since they are poikilothermic species that need sunlight to regulate their internal temperature, lizard frequent lower altitudes in search of clearings that may appear due to forestry harvesting (Delgado and Ferro, 2006).
Illumination
Weighting: frequency by illumination
The highest use of this resource was in the category established as sun filtered by vegetation (Figure 3), whose frequency was stable throughout all the years of study for the two treatments. In the five-year post-logging treatment, the values of filtered sun use had the greatest variability.
With the effect of locality, the categories of illumination of sun filtered by vegetation and sun and year were significant. As for the treatments, no significant associations were shown, the season indicated significant associations for the three established categories of illumination, but not with the years. The other associations in the univariate analysis were not significant for 5 %. In the multivariate analysis, only the "Location*treatment" combination was significant when controlling for lighting strata with an advantage ratio of 0.910 and an advantage ratio homogeneity significance of 0.007.
In both locations and for the intermediate year, there was a significant decrease in the use of the sunny category and an increase in the shade category. At Cabo de San Antonio there was no defined pattern of these changes between treatments, but there was at Cabo Corrientes, where the use of the sunny category was greater in treatment 1, while the shade category was greater in treatment 2.
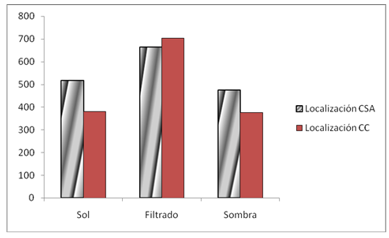
Fig. 3 - Lizard frequency. Location Cabo de San Antonio (CSA) and Cabo Corrientes (CC) by the three established categories of illumination
The behavior of the lighting resource was stable in all years of study, always preferring sites with filtered sun. As mentioned in previous analyses, it is a vertebrate group that does not regulate its internal temperature and prolonged periods of exposure to direct sunlight, as well as total shade, can affect the proper functioning of the organism and the performance of its vital functions (Rodríguez Shettino 1999).
The cost of thermoregulation depends largely on the habitat: raising body temperature is more difficult in closed forests than in more open areas, so that species in environments costly for thermoregulation are generally passive to changes in ambient temperature, i.e., they are thermoconformable (Huey y Slatkin 1976).
In both locations, in 2013, there was a notable decrease in the use of the direct sun category and an increase in the shade category, the differences are not significant, but it is assumed that the climatic conditions of this year were different from the rest of the years, the rainy period was above the historical norm, as mentioned above, and the low rainfall period was shorter, so the expiration period of the trees decreased; This type of forest is identified by having between 40 and 70 % of the trees in the upper tree stratum lose their leaves during the low rainfall period (Del Risco 1999) and the shorter period reduces this effect, which means that these changes have had a differential influence on the use of the resources discussed here by the lizard community.
In Cabo Corrientes the use of the direct sun category was higher in treatment 1. The vegetation structure conditions the ambient temperature of each type of habitat, but it also underlies different possibilities of vertically occupying the space, the treatment of 5 years of post-logging recovery has the lowest vegetation cover and consequently the highest solar incidence, while those of more than 30 years without intervention have a higher incidence of shade.
Substrate
Weighting: frequency by substrate
In the analysis of substrate use in both locations, the most used for all combinations (years, treatments and seasons) were tree trunks (Figure 4) with a tendency to increase their use in the treatment of more than 30 years post logging.
In this case, the treatment*substrate association was significant in the branch/soil category, the number of times that lizards were more likely to be found on the branches than on the ground was 6.98. The season*substrate association showed a significant difference in the use of substrates. The multivariate analysis gave a very high Chi-square value indicating a lack of homogeneity in the advantage ratio between substrates in the combination "Location* Season".
Throughout the three years of the study, there were no statistically significant changes in the use of the log resource, but there was a tendency to increase its use in the treatment of more than 30 years after logging, with a range of values between 40 and 70 %. The use of this resource increased in the last two years of the study equally in both treatments.
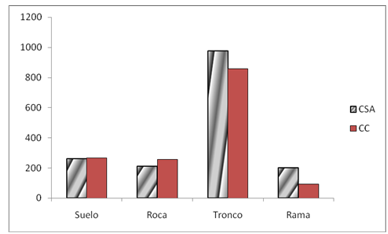
Fig. 4 - Frequency of lizards by locality Cabo de San Antonio (CSA) and Cabo Corrientes (CC) by the four established substrate categories
In all localities and periods, a preference for logs as substrate was observed. Similar results have been found by Rodriguez Schettino (1999), coinciding in the selection of the group. The most abundant species in the community are sympatric species of the trunk-soil ecomorph.
Forest harvesting is characterized by the extraction of individuals for different purposes, and when high volumes of wood are extracted from a plant formation, its structure is modified and brings with it changes such as increased temperature, decreased humidity, changes in light and wind speed (Pinard and Putz, 1996), but in the following years the natural response of the forest leads to a recolonization of these spaces by invasive, opportunistic and austere species (Delgado et al., 2005), which increases plant density, and with the passage of time, due to competition between all the new individuals and the remaining ones, the less adapted ones are eliminated, and therefore the number of individuals per hectare decreases.
It should also be considered that the most preserved forests have among their characteristics a greater roughness of the trunks by the species that characterize it (Ferro, 2004), and this favors the presence of a great variety of microhabitats for invertebrates and their accumulation, which serves as food for the group under analysis.
The particularities appear in the Cabo de San Antonio locality where the use of branches increased in the rainy period, this period was the rainiest of the three years sampled with values above the historical average (1 480 mm) and relative humidity increased, moving individuals to higher altitudes.
The explanation for these results is that the survival and reproduction of lizards is influenced by the use of the soil, perches or structural niche and the availability of food or trophic niche (Shoener, 1968), elements that can be altered in such a way that forest management favors some species to the detriment of others.
When correlating the densities of the vegetation strata and the preference of the four categories of substrates established for this study, there are significant negative results between the preference of rocks and branches with the shrub stratum. It can be assumed that this is due to the fact that when rock density decreases, the density of the shrub stratum increases, and as the density of this stratum is higher, branching is lower, since it increases in the less dense areas.
Relationship with vegetation variables. Study of the strength of the relationship between the potential predictor variables and the dependent variable.
The total frequency of lizards correlated significantly with the richness of the shrub stratum (-0.294), height of the upper tree stratum (0.208), mean length of fallen logs (0.182) and natural regeneration (-0.261). The frequency of males only showed significant relationship with the richness of the shrub layer (-0.206) and natural regeneration (-0.189) as well as adults (-0.230 and -0.189 respectively).
Figures 5 and 6 present the results of the canonical correspondence analyses carried out between the vegetation variables of each study treatment and the frequencies of lizard species in each of them. It is found that the time of forest recovery influences the differences between lizard associations with these variables. Anolis quadriocellifer is the species that is not affected by the vegetation variables measured in the two study treatments in the Cabo de San Antonio sector.
In the analysis of the rainy period, the first quadrant of Figure 5 shows the species that preferentially use the soil and rocks as substrate, Anolis loysiana and Leiocephalus macropus are associated with the basal area of the vegetation formation, since their density was maximum in the plots with the highest numbers.
Leiocephalus carinatus frequented sites with higher shrub stratum heights, while Ameiva auberi was more detected in areas of high volume. In the second quadrant it is only observed two species of lizards, Anolis porcatus and A. homolechis associated with the richness of the trees and sites of higher densities respectively. On the other hand, Anolis angusticeps and A. alutaceu showed no association with any of the vegetation variables measured. The first species was always more detected in open spaces with sun penetration and in treatments with less time of forest harvesting, it was found in the branch and leaf substrate in equal proportion, because the movements of this small lizard are slow and it has the habit of staying very attached to the branches (Böhm et al., 2013); however, few individuals of Anolis alutaceu were observed, mostly on the ground and in the shade, coinciding with what was stated by Schwartz and Henderson (1991).
In the low rainfall period, the behavior of the species differs from the previous one, except in the case of the regional endemic, which is the only species that occupies the four types of substrate in both areas with a high frequency on the trunk of trees from 70 % to 80 %. (Pérez et al., 2014). In the first quadrant the variables of vegetation volume and basal area in association with Anolis homolechis, A. porcatus, A. luteogularis, Leiocepahaulus macropus, L. carinatus and Ameiva auberi, were recorded in plots with greater specific richness of vegetation, where a segregation strategy to reduce competition for resources is evident, coinciding these results with those of González (2017), while Anolis alutaceus and Anolis loysiana showed no association with any measured variable.
 Note: Vegetation variables are shown as arrows, census units are shown as circles and working treatments as squares. Aq, Anolis quadriocellifer*, Ah, Anolis homolechis*, Al, Anolis loysiana*, Aan, Anolis angusticeps, Ap, Anolis porcatus*, As, Anolis sagrei, Lc, Leiocephalus carinatus, Lm, Leiocephalus macropus*, Ama, Ameiva auberi, Aall, Anolis allogus*, Aal, Anolis alutaceus*, Alut, Anolis luteogularis*, Ls, Leiocephalus stictigaster*, G ha-1, Basal area, hma, Mean shrub layer height, hmai, Mean lower tree layer height, V ha-1, Volume, Da, Density of shrub layer, Dai, Density of lower tree layer, Sp arbr, Species richness, Nreg, Natural regeneration.
Note: Vegetation variables are shown as arrows, census units are shown as circles and working treatments as squares. Aq, Anolis quadriocellifer*, Ah, Anolis homolechis*, Al, Anolis loysiana*, Aan, Anolis angusticeps, Ap, Anolis porcatus*, As, Anolis sagrei, Lc, Leiocephalus carinatus, Lm, Leiocephalus macropus*, Ama, Ameiva auberi, Aall, Anolis allogus*, Aal, Anolis alutaceus*, Alut, Anolis luteogularis*, Ls, Leiocephalus stictigaster*, G ha-1, Basal area, hma, Mean shrub layer height, hmai, Mean lower tree layer height, V ha-1, Volume, Da, Density of shrub layer, Dai, Density of lower tree layer, Sp arbr, Species richness, Nreg, Natural regeneration.Fig. 5 - Ordination diagram of correspondence analysis performed with the density of lizard species and vegetation variables in the Cabo de San Antonio sector
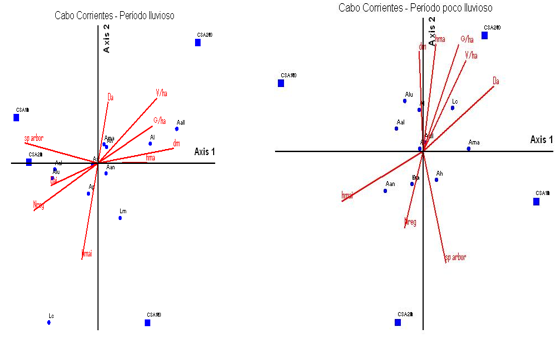
In the Cabo Corrientes locality, the species that showed the least modification with the vegetation variables was Anolis sagrei, which did not share its ecological niche in either of the two periods with Anolis quadriocellifer. In the analysis of the rainy period, in the second quadrant, Anolis homolechis and Ameiva auberi were associated with the variables of vegetation, volume and density of the lower tree layer, while Anolis loysina and Anolis allogus were associated with the highest values of basal area and tree diameter, in the treatments of more than 30 years of post-logging recovery.
Leiocephalus macropus in this period is far from the rest of the species, while in the low rainfall period it preferred the habitats where there is a greater natural regeneration, which coincides with treatments with a shorter post-harvesting recovery time (Uvero Quemado).
Anolis porcatus was associated with higher altitudes and Anolis luteogularis with higher densities. Anolis homolechis had a different behavior in this period compared to the previous one, since in this case its selected habitats with higher species richness.
The species that resulted away from the others were Ameiva auberi and A. alutaceus, these two species have different uses of the resource, the first was observed occupying the soil substrate and rocks, exposed to direct sun or filtered by vegetation, the other species preferred low altitudes and shade.
Anolis allogus overlaps with Anolis sagrei preferring sunny forest sites and higher altitudes.
In the low rainfall period of this locality, Anolis allogus and Leiocephalus carinatus are associated with plots with higher tree volumes, basal area and heights, Ameiva auberi with the densest sites, Anolis luteogularis and A. alutaceus show no association with any measured variable and Anolis angusticeps with the highest heights.
Anolis loysina and Anolis allogus are associated with the highest values of basal area and diameter of the trees in the treatments of more than 30 years of post-logging recovery, but it is this recovery time that leads to the plant formation being characterized by the increase of these indicators, because after the competition and elimination of the less adapted individuals, the remaining ones have larger spaces, more homogeneously distributed and thus develop larger diameters and basal areas (Delgado and Ferro 2006).
Anolis porcatus and Anolis angusticeps are associated with plots with higher altitudes and Anolis luteogularis with higher densities, all of them are species that frequent the highest substrates in the plant formation (trunks and branches).
In the low rainfall period, Anolis allogus and Leiocephalus carinatus were associated with plots with higher tree volumes, basal areas and heights, while Ameiva auberi frequented the densest sites with the highest heights.
Lizards may maintain viable populations in isolated fragments for longer periods than birds and mammals (Hinsley et al., 1996; Henle et al., 2004), ensuring their persistence in fragmented landscapes despite their reduced colonization ability (Hinsley et al., 1996; Henle et al., 2004).
 Note: Vegetation variables are shown as arrows, species are shown as circles and working treatments as squares. Aq, Anolis quadriocellifer*, Ah, Anolis homolechis*, Al, Anolis loysiana*, Aan, Anolis angusticeps, Ap, Anolis porcatus*, As, Anolis sagrei, Lc, Leiocephalus carinatus, Lm, Leiocephalus macropus*, Ama, Ameiva auberi, Aall, Anolis allogus*, Aal, Anolis alutaceus*, Alut, Anolis luteogularis*, Ls, Leiocephalus stictigaster*, G ha-1, Basal area, hma, Mean shrub layer height, hmai, Mean lower tree layer height, V ha-1, Volume, Da, Density of shrub layer, Dai, Density of lower tree layer, Sp arbr, Species richness, Nreg, Natural regeneration.
Note: Vegetation variables are shown as arrows, species are shown as circles and working treatments as squares. Aq, Anolis quadriocellifer*, Ah, Anolis homolechis*, Al, Anolis loysiana*, Aan, Anolis angusticeps, Ap, Anolis porcatus*, As, Anolis sagrei, Lc, Leiocephalus carinatus, Lm, Leiocephalus macropus*, Ama, Ameiva auberi, Aall, Anolis allogus*, Aal, Anolis alutaceus*, Alut, Anolis luteogularis*, Ls, Leiocephalus stictigaster*, G ha-1, Basal area, hma, Mean shrub layer height, hmai, Mean lower tree layer height, V ha-1, Volume, Da, Density of shrub layer, Dai, Density of lower tree layer, Sp arbr, Species richness, Nreg, Natural regeneration.Fig. 6 - Ordination diagram of correspondence analysis performed with the density of lizard species and vegetation variables in the Cabo de San Antonio sector. Cape Corrientes
The Mantel test showed that there is a correlation between the vegetation variables in the two locations studied (r= 0.86; P < 0.05) and the densities of lizards. According to the above results, it can be established that there are large variations in the relationships of the lizard communities with the vegetation structure between rainy and low rainfall periods.
The result of the canonical correspondence analysis is due to microhabitat selection as shown in Figure 5 and Figure 6 could be a mechanism explaining the persistence of lizard populations in relation to associated vegetation variables and post-harvest recovery time of the forests under study, since microhabitat selection can vary in space and time with respect to food sources, reproduction, predators and other habitat quality components (Martín and López 2002). Agreeing with Caro (2010), in his review of animal groups that can be indicators of various anthropogenic and natural disturbances, he points out that reptiles could serve as indicators of forest management.
With this study, it has been possible to point out the relationship of each species of lizard with the different elements of vegetation and the recovery time after logging for each study locality, thus defining the indicators for each structural feature and period of reestablishment.
The species of this genus, with a wide ecological niche, are more likely to make ecological displacements for some of the habitat dimensions, in search of shelter, food and minimum survival conditions. The study of lizards has recently focused on determining the effects that forest management can have on displacement due to vegetation alterations (Arias and Martínez 2014). This is intended to demonstrate that lizards could be indicators of forest management disturbances and other anthropogenic impacts (Hager 1998).
Lizard species richness and vegetation richness associated with distribution range limits have been studied by different authors (Schall and Pianka 1977, Bock et al., 1981 and Uetz 2000). However, these analyses do not reveal the ultimate causes of distribution patterns of ecological mechanisms (Rojas et al., 2016). Other research has focused on the habitat preferences of particular species or communities, sometimes considering the relationships between habitat features and differential performance of individuals or populations, growth rate and survival (Medina and Cardena 2015).
CONCLUSIONS
The use of resources by the detected species varied significantly with the time of post-logging recovery in the study localities for height and illumination. In addition, the period of the year had an influence on this indicator, with the period of low rainfall showing less variation in the preference of the established categories in the use of the resource.
The vegetation variables that showed the greatest association with the different species of lizards were species richness, density of the lower tree stratum and height of the three established strata.
The associations established between changes in forest structure and lizard communities are significantly controlled by the height and illumination strata in both locations and the vegetation variables that have a greater correspondence are richness of the shrub stratum, height of the upper tree stratum and natural regeneration.
REFERENCIAS BIBLIOGRÁFICAS
ALBERTS, A.C., LEMM, J.M., PERRY, A.M., MORICI, L.A. y PHILLIPS, J.A., 2002. Temporary alteration of local social structure in a threatened population of Cuban iguanas (Cyclura nubila). Behavioral Ecology and Sociobiology [en línea], vol. 51, no. 4, pp. 324-335. [Consulta: 02/03/2021]. ISSN 1432-0762. DOI 10.1007/s00265-001-0445-z. Disponible en: Disponible en: https://doi.org/10.1007/s00265-001-0445-z . [ Links ]
ARIAS, B.A. y MARTÍNEZ, R.M., 2014. Comunidades de lagartos terrestres. Fauna terrestre del Archipiélago de Sabana-Camagüey [en línea]. Cuba: Sello Editorial Academia, pp. 188-192. ISBN 978-959-270-294-3. Disponible en: https://www.semanticscholar.org/paper/Fauna-terrestre-del-Archipi%C3%A9lago-de-Cuba-Batista-Barreto/1da5a37c49dccaa3d9abf25ade5cc4d9caa2f85b. [ Links ]
BOCK, J., BOCK, C. y FRITZ, R., 1981. Biogeography of Illinois Reptiles and Amphibians: A Numerical Analysis. American Midland Naturalist [en línea], DOI 10.2307/2425162. Disponible en: https://www.semanticscholar.org/paper/Biogeography-of-Illinois-Reptiles-and-Amphibians%3A-A-Bock-Bock/efc08599f0a6605f9a9f588daca68d60da1c4a50. [ Links ]
BÖHM, M., et al., 2013. The conservation status of the world’s reptiles. Biological Conservation [en línea], vol. 157, pp. 372-385. [Consulta: 02/03/2021]. ISSN 0006-3207. DOI 10.1016/j.biocon.2012.07.015. Disponible en: Disponible en: https://www.sciencedirect.com/science/article/pii/S0006320712003357 . [ Links ]
CARO, T., 2010. Conservation by Proxy: Indicator, Umbrella, Keystone, Flagship, and Other Surrogate Species [en línea]. S.l.: Island Press. ISBN 978-1-59726-959-9. Disponible en: https://books.google.com.cu/books/about/Conservation_by_Proxy.html?id=i0W5yf4L12EC&redir_esc=y. [ Links ]
COGOLLO, J. y URBINA-CARDONA, N., 2008. Patrones de Diversidad y Composición de Reptiles en Fragmentos de Bosque Seco Tropical en Córdoba, Colombia. Tropical Conservation Science [en línea], vol. 1, pp. 397-416. DOI 10.1177/194008290800100407. Disponible en: https://www.researchgate.net/publication/216019496_Patrones_de_Diversidad_y_Composicion_de_Reptiles_en_Fragmentos_de_Bosque_Seco_Tropical_en_Cordoba_Colombia. [ Links ]
DELGADO, F. y FERRO, J., 2000. La regeneración natural de bosques semideciduos en la Reserva de Biosfera Península de Guanahacabibes. La Habana, Cuba: Agencia de Ciencia y Tecnología, CITMA. Informe final de Resultado Parcial Proyecto 01302079 PNCT «Los Cambios Globales y la Evolución del Medio Ambiente en Cuba». [ Links ]
DELGADO, F. y FERRO, J., 2006. Productividad y mortalidad de los bosques semideciduos de a RBPG. Memorias del X Congreso Latinoamericano de Botánica. República Dominicana: s.n., ISBN 9945-428-00-4. [ Links ]
DUELLMAN, W.E., 1979. The South American herpetofauna: its origin, evolution, and dispersal [en línea]. Lawrence, Kan.: Museum of Natural History, University of Kansas. ISBN 978-0-89338-008-3. Disponible en: https://www.worldcat.org/title/south-american-herpetofauna-its-origin-evolution-and-dispersal/oclc/6212016. [ Links ]
FERNÁNDEZ, L.H., FERNÁNDEZ, F.D. y DÍAZ, J.F., 2005. Capacidad competitiva de las especies forestales de los bosques semideciduos en la Reserva de la Biosfera Península de Guanahacabibes. Mapping [en línea], no. 100, pp. 44-49. [Consulta: 02/03/2021]. ISSN 1131-9100. Disponible en: Disponible en: https://dialnet.unirioja.es/servlet/articulo?codigo=1112790 . [ Links ]
FERRO, J., 2004. Efecto del aprovechamiento forestal sobre la estructura y dinámica de la comunidad de epifitas vasculares del bosque semideciduonotófilo de la Península de Guanahacabibes. Tesis en opción al grado científico Doctor en Ciencias Forestales. Pinar del Río, Cuba: Universidad de Pinar del Río «Hermanos Saíz Montes de Oca». [ Links ]
GONZÁLEZ PÉREZ, D., 2017. Patrones de segregación de la comunidad de lagartos terrestres diurnos en cayo Paredón Grande, archipiélago Sabana-Camagüey, Cuba [en línea]. Thesis. Universidad Central “Marta Abreu” de Las Villas. Facultad de Ciencias Agropecuarias. Departamento de Biología. [Consulta: 02/03/2021]. Disponible en: Disponible en: http://dspace.uclv.edu.cu:8089/xmlui/handle/123456789/7908 . [ Links ]
HAGER, H.A., 1998. Area-sensitivity of reptiles and amphibians: Are there indicator species for habitat fragmentation? Écoscience [en línea], vol. 5, no. 2, pp. 139-147. [Consulta: 02/03/2021]. ISSN 1195-6860. DOI 10.1080/11956860.1998.11682463. Disponible en: Disponible en: https://doi.org/10.1080/11956860.1998.11682463 . [ Links ]
HENLE, K., LINDENMAYER, D.B., MARGULES, C.R., SAUNDERS, D.A. y WISSEL, C., 2004. Species Survival in Fragmented Landscapes: Where are We Now? Biodiversity & Conservation [en línea], vol. 13, no. 1, pp. 1-8. [Consulta: 02/03/2021]. ISSN 1572-9710. DOI 10.1023/B:BIOC.0000004311.04226.29. Disponible en: Disponible en: https://doi.org/10.1023/B:BIOC.0000004311.04226.29 . [ Links ]
HINSLEY, S.A., BELLAMY, P.E., NEWTON, I. y SPARKS, T.H., 1996. Influences of population size and woodland area on bird species distributions in small woods. Oecologia [en línea], vol. 105, no. 1, pp. 100-106. [Consulta: 02/03/2021]. ISSN 1432-1939. DOI 10.1007/BF00328797. Disponible en: Disponible en: https://doi.org/10.1007/BF00328797 . [ Links ]
HUEY, R.B. y SLATKIN, M., 1976. Cost and benefits of lizard thermoregulation. The Quarterly Review of Biology, vol. 51, no. 3, pp. 363-384. ISSN 0033-5770. DOI 10.1086/409470. [ Links ]
LAURIE, J.V. y R. PIANKA, E., 2005. Deep history impacts present-day ecology and biodiversity. Localización: Proceedings of the National Academy of Sciences of the United States , no. 22. Disponible en: https://dialnet.unirioja.es/servlet/articulo?codigo=1189311. [ Links ]
LOSOS, J., 2009. Lizards in an Evolutionary Tree: Ecology and Adaptive Radiation of Anoles [en línea]. S.l.: University of California Press. ISBN 978-0-520-94373-5. Disponible en: https://books.google.com.cu/books/about/Lizards_in_an_Evolutionary_Tree.html?id=fyu6xlGnsc0C&redir_esc=y. [ Links ]
MARTÍN, J. y LOPEZ, P., 2002. The effect of Mediterranean dehesa management on lizard distribution and conservation. Biological Conservation [en línea], vol. 108, no. 2, pp. 213-219. [Consulta: 02/03/2021]. ISSN 0006-3207. DOI 10.1016/S0006-3207(02)00107-6. Disponible en: Disponible en: https://www.sciencedirect.com/science/article/pii/S0006320702001076 . [ Links ]
MEDINA-RANGEL, G.F., CÁRDENAS-ÁREVALO, G., MEDINA-RANGEL, G.F. y CÁRDENAS-ÁREVALO, G., 2015. Relaciones espaciales y alimenticias del ensamblaje de reptiles del complejo cenagoso de Zapatosa, departamento del Cesar (Colombia). Papéis Avulsos de Zoologia [en línea], vol. 55, no. 10, pp. 143-165. [Consulta: 02/03/2021]. ISSN 0031-1049. DOI 10.1590/0031-1049.2015.55.10. Disponible en: Disponible en: http://www.scielo.br/scielo.php?script=sci_abstract&pid=S0031-10492015001000143&lng=en&nrm=iso&tlng=es . [ Links ]
MESQUITA, D.O., COSTA, G.C. y COLLI, G.R., 2006. ECOLOGY OF AN AMAZONIAN SAVANNA LIZARD ASSEMBLAGE IN MONTE ALEGRE, PARÁ STATE, BRAZIL. South American Journal of Herpetology [en línea], vol. 1, no. 1, pp. 61-71. [Consulta: 02/03/2021]. ISSN 1808-9798, 1982-355X. DOI 10.2994/1808-9798(2006)1[61:EOAASL]2.0.CO;2. Disponible en: Disponible en: https://bioone.org/journals/south-american-journal-of-herpetology/volume-1/issue-1/1808-9798(2006)1[61:EOAASL]2.0.CO;2/ECOLOGY-OF-AN-AMAZONIAN-SAVANNA-LIZARD-ASSEMBLAGE-IN-MONTE-ALEGRE/10.2994/1808-9798(2006)1[61:EOAASL]2.0.CO;2.full . [ Links ]
MURCIA, L.E.R., COGOLLO, J.E.C. y BELLO, J.A.C., 2016. Reptiles del bosque seco estacional en el Caribe colombiano: distribución de los hábitats y del recurso alimentario. Acta biológica colombiana [en línea], vol. 21, no. 2, pp. 365-377. [Consulta: 02/03/2021]. ISSN 0120-548X. Disponible en: Disponible en: https://dialnet.unirioja.es/servlet/articulo?codigo=5974699 . [ Links ]
PÉREZ RODRÍGUEZ, E., BEROVIDES, V. y DELGADO, F., 2014. Abundancia y diversidad de comunidades de lagartos en el sector más oriental del Parque Nacional Guanahacabibes. Revista Científica Cuba Zoo [en línea], no. 25, pp. 34. [Consulta: 02/03/2021]. Disponible en: Disponible en: http://revista.cubazoo.cu/archive/edicion25-2014/Abundancia-y-diversidad-de-comunidades-de-lagartos-en-el-sector-mas-oriental-del-PZN-Ganahacabibes.pdf . [ Links ]
PINARD, M.A. y PUTZ, F.E., 1996. Conserving forest biomass by reducing logging damage. Biotropica, no. 28, pp. 278-95. [ Links ]
RAND, A.S., 1964. Ecological Distribution in Anoline Lizards of Puerto Rico. Ecology [en línea], vol. 45, no. 4, pp. 745-752. [Consulta: 02/03/2021]. ISSN 1939-9170. DOI 10.2307/1934922. Disponible en: Disponible en: https://esajournals.onlinelibrary.wiley.com/doi/abs/10.2307/1934922 . [ Links ]
RODRÍGUEZ, E. del R., 1999. Cuban Forests: Their History and Characteristics [en línea]. S.l.: Editorial José Martí. ISBN 978-959-09-0160-7. Disponible en: https://books.google.com.cu/books/about/Cuban_Forests.html?id=HCYkAAAACAAJ&redir_esc=y. [ Links ]
RODRÍGUEZ SCHETTINO, L. y COY OTERO, A., 1999. The iguanid lizards of Cuba [en línea]. Gainesville: University Press of Florida. ISBN 978-0-8130-1647-4. Disponible en: https://www.worldcat.org/title/iguanid-lizards-of-cuba/oclc/40588341. [ Links ]
SCHALL, J.J. y PIANKA, E.R., 1977. Species densities of reptiles and amphibians on the Iberian península. Donaria, Acta Vertehr.ata, no. 4, pp. 27-34. [ Links ]
SCHETTINO, L.R., MANCINA, C.A. y GONZÁLEZ, V.R., 2013. Reptiles of Cuba: Checklist and Geographic Distributions [en línea]. S.l.: Verlag nicht ermittelbar. Disponible en: https://books.google.com.cu/books/about/Reptiles_of_Cuba.html?id=EAqkoAEACAAJ&redir_esc=y. [ Links ]
SCHOENER, T.W. y GORMAN, G.C., 1968. Some Niche Differences in Three Lesser Antillean Lizards of the Genus Anolis. Ecology [en línea], vol. 49, no. 5, pp. 819-830. [Consulta: 02/03/2021]. ISSN 1939-9170. DOI 10.2307/1936533. Disponible en: Disponible en: https://esajournals.onlinelibrary.wiley.com/doi/abs/10.2307/1936533 . [ Links ]
SCHWARTZ, A. y HENDERSON, R.W., 1991. Amphibians and Reptiles of the West Indies: Descriptions, Distributions, and Natural History [en línea]. S.l.: University of Florida Press. ISBN 978-0-8130-1049-6. Disponible en: https://books.google.com.cu/books/about/Amphibians_and_Reptiles_of_the_West_Indi.html?id=0x_hxEolYUAC&redir_esc=y. [ Links ]
Received: November 29, 2020; Accepted: March 01, 2021

